
Deux les gènes sont liés quand ils ont tendance à être hérités ensemble comme s'ils formaient une seule entité. Cela peut également se produire avec plus de deux gènes. Dans tous les cas, ce comportement des gènes est ce qui a permis la cartographie génétique par liaison et recombinaison..
À l'époque de Mendel, d'autres chercheurs, tels que les époux Boveri, avaient observé qu'il y avait des corps dans le noyau de la cellule qui sécrétaient pendant le processus de division cellulaire. C'étaient les chromosomes.

Plus tard, avec les travaux de Morgan et de son groupe, il y avait une meilleure compréhension de l'hérédité des gènes et des chromosomes. Autrement dit, les gènes se séparent comme les chromosomes qui les portent (théorie chromosomique de l'hérédité).
Index des articles
Il y a, comme nous le savons, beaucoup moins de chromosomes que de gènes. L'être humain, par exemple, possède environ 20000 gènes répartis dans environ 23 chromosomes différents (charge haploïde de l'espèce).
Chaque chromosome est représenté par une longue molécule d'ADN dans laquelle de très nombreux gènes sont codés séparément. Chaque gène réside donc sur un site particulier (locus) sur un chromosome spécifique; à son tour, chaque chromosome porte de nombreux gènes.
En d'autres termes, tous les gènes d'un chromosome sont liés les uns aux autres. S'il semble que non, c'est parce qu'il y a un processus d'échange physique d'ADN entre les chromosomes qui crée l'illusion d'une distribution indépendante..
Ce processus s'appelle la recombinaison. Si deux gènes sont liés, mais largement séparés l'un de l'autre, la recombinaison se produira toujours et les gènes se sépareront comme Mendel l'a observé..
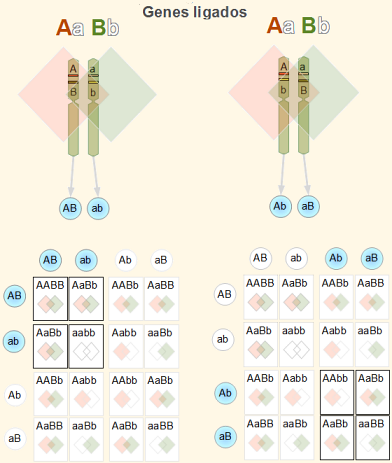
Pour observer et démontrer le lien, le chercheur procède à des croisements avec des individus qui présentent une manifestation contrastée du phénotype des gènes à l'étude (par exemple, P: AAbb X aaBB).
Tous les descendants de F1 seront AaBb. De la croix dihybride AaBb X aabb (ou test croisé) on s'attendrait à une progéniture F2 présentant les proportions génotypiques (et phénotypiques) 1AaBb:1Aabb:1aaBb:1aabb.
Mais cela n'est vrai que si les gènes ne sont pas liés. La première indication génétique que deux gènes sont liés est qu'une prédominance des phénotypes paternels est observée: c'est-à-dire, Aabb + aaBb >> AaB_b + aabb.
Dans le cas des gènes liés que nous utilisons à titre d'exemple, les individus produiront principalement des gamètes Un B Oui un B, plus que des gamètes UN B Oui un B.
Puisque l'allèle dominant d'un gène est associé à l'allèle récessif de l'autre gène, on dit que les deux gènes sont liés dans la répulsion. Si une prédominance d'allèles est observée UN B Oui un B à propos des gamètes Un B Oui un B, on dit que les gènes sont liés dans le couplage.
Autrement dit, les allèles dominants sont couplés à la même molécule d'ADN; ou ce qui est pareil, ils sont associés au même chromosome. Cette information est extrêmement utile dans l'amélioration génétique.
Cela permet d'établir le nombre d'individus à analyser lorsque les gènes sont liés et on souhaite sélectionner, par exemple, les deux caractères dominants.
Cela serait plus difficile à réaliser lorsque les deux gènes sont en répulsion et que la liaison est si étroite qu'il n'y a pratiquement pas de recombinaison entre les deux gènes..
L'existence de la liaison elle-même a été une énorme avancée dans notre compréhension des gènes et de leur organisation. Mais en plus, cela nous a aussi permis de comprendre comment la sélection peut agir dans les populations et expliquer un peu l'évolution des êtres vivants.
Il y a des gènes qui sont si étroitement liés que seuls deux types de gamètes sont produits au lieu des quatre qui permettraient une distribution indépendante.
Dans les cas extrêmes, ces deux gènes liés (en couplage ou en répulsion) n'apparaissent que dans un type d'association dans la population. Si cela se produit, on dit qu'il existe un déséquilibre de liaison.
Le déséquilibre de liaison se produit, par exemple, lorsque l'absence des deux allèles dominants diminue les chances de survie et de reproduction des individus..
Cela se produit lorsque les individus sont le produit de la fécondation entre gamètes un B. Fécondation entre gamètes un B Oui Un B, au contraire, il augmente la probabilité de survie de l'individu.
Ceux-ci auront au moins un allèle À et un allèle B, et affichera les fonctions associées sauvages correspondantes.
La liaison et son déséquilibre peuvent également expliquer pourquoi certains allèles indésirables d'un gène ne sont pas éliminés de la population. S'ils sont étroitement liés (en répulsion) aux allèles dominants d'un autre gène qui confère des avantages à leur porteur (par exemple, un B), être associé au «bon» permet la permanence du «mauvais».
Une conséquence importante de la liaison est qu'elle permet de déterminer la distance entre les gènes liés. Cela s'est avéré être historiquement vrai et a conduit à la génération des premières cartes génétiques..
Pour cela, il était nécessaire de comprendre que les chromosomes homologues peuvent se croiser au cours de la méiose dans un processus appelé recombinaison..
Lors de la recombinaison, des gamètes différents sont produits par rapport à ceux qu'un individu ne pourrait produire que par ségrégation. Puisque les recombinants peuvent être comptés, il est possible d'exprimer mathématiquement la distance entre un gène et un autre.
Dans les cartes de liaison et de recombinaison, les individus qui sont recombinés entre une paire de gènes particulière sont comptés. Ensuite, son pourcentage est calculé en fonction de la population cartographique totale employée..
Par convention, un pour cent (1%) de recombinaison est une unité de carte génétique (umg). Par exemple, dans une population cartographique de 1000 individus, 200 recombinants sont trouvés parmi les marqueurs génétiques. À/à Oui B/b. Par conséquent, la distance qui les sépare sur le chromosome est de 20 umg.
Actuellement, 1 umg (soit 1% de recombinaison) est appelé cM (centi Morgan). Dans le cas ci-dessus, la distance entre À/à Oui B/b est de 20 cM.
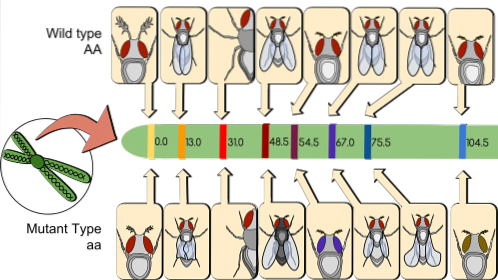
Dans une carte génétique, les distances en cM peuvent être ajoutées, mais il est évident que les pourcentages de recombinaison ne peuvent pas être ajoutés. Les gènes doivent toujours être cartographiés suffisamment éloignés pour pouvoir mesurer de courtes distances.
Si la distance entre deux marqueurs est très élevée, la probabilité qu'il y ait un événement de recombinaison entre eux est égale à 1. Par conséquent, ils se recombineront toujours et ces gènes se comporteront comme s'ils étaient distribués indépendamment même s'ils sont liés..
En revanche, pour diverses raisons, les cartes mesurées en cM ne sont pas linéairement liées à la quantité d'ADN impliquée. De plus, la quantité d'ADN par cM n'est pas universelle, et pour chaque espèce particulière, il s'agit d'une valeur particulière et moyenne..



Personne n'a encore commenté ce post.