Avec "héritage non mendélien«Nous entendons tout modèle d'héritage dans lequel les personnages hérités ne se séparent pas conformément aux dispositions des lois de Mendel..
En 1865, Gregor Mendel, considéré comme le «père de la génétique», effectue une série de croisements expérimentaux avec des plants de pois dont les résultats le conduisent à proposer des postulats (lois de Mendel) qui cherchent à donner une explication logique à l'hérédité. les personnages entre parents et enfants.
Cet astucieux moine autrichien a soigneusement observé la ségrégation des gènes parentaux et leur apparition chez la progéniture en tant que caractères dominants et récessifs. De plus, il a déterminé les modèles mathématiques qui décrivaient l'héritage d'une génération à l'autre et ces résultats ont été «ordonnés» sous la forme de 3 lois fondamentales:
- La loi de la domination
- La loi de la ségrégation des personnages et
- La loi de la distribution indépendante.
Les succès et déductions de Mendel ont été cachés pendant de nombreuses années, jusqu'à leur redécouverte au début du XXe siècle..

À cette époque, cependant, la communauté scientifique a maintenu une position quelque peu sceptique à l'égard de ces lois, car elles ne semblaient pas expliquer les modèles d'héritabilité dans aucune espèce animale ou végétale, en particulier dans les caractères déterminés par plus d'un locus..
De ce fait, les premiers généticiens ont classé les patrons héréditaires observés comme «mendéliens» (ceux qui pourraient s'expliquer par la ségrégation d'allèles simples, dominants ou récessifs, appartenant au même locus) et «non mendéliens» (ceux qui ne pourrait être expliqué si facilement).
Index des articles
L'hérédité mendélienne fait référence à un schéma héréditaire qui respecte les lois de la ségrégation et de la distribution indépendante, selon lequel un gène hérité de n'importe quel parent sécrète dans des gamètes avec une fréquence équivalente ou, plutôt, avec la même probabilité.
Les principaux modèles d'hérédité mendélienne qui ont été décrits pour certaines maladies sont: autosomique récessif, autosomique dominant et lié au chromosome X, qui s'ajoutent aux modèles de dominance et de récessivité décrits par Mendel.
Cependant, ceux-ci ont été postulés concernant les traits visibles et non les gènes (il convient de noter que certains allèles peuvent coder des traits qui se séparent en tant que dominants, tandis que d'autres peuvent coder les mêmes traits, mais ceux-ci se séparent en tant que gènes récessifs).
De ce qui précède, il s'ensuit que l'héritage non mendélien consiste simplement en tout modèle héréditaire qui ne répond pas à la norme dans laquelle un gène hérité d'un parent sécrète dans des cellules germinales avec les mêmes probabilités, et ceux-ci incluent:
- Héritage mitochondrial
- "Impression"
- Disomie uniparentale
- Dominance incomplète
- Codominance
- Allèles multiples
- Pléiotropie
- Allèles létaux
- Traits polygéniques
- Héritage lié au sexe
L'apparition de ces variations dans les modèles héréditaires peut être attribuée aux diverses interactions que les gènes ont avec d'autres composants cellulaires, en plus du fait que chacun est soumis à une régulation et à une variation à l'un des stades de la transcription, de l'épissage, de la traduction, des protéines. pliage, oligomérisation, translocation et compartimentation au sein de la cellule et pour son exportation.
En d'autres termes, il existe de nombreuses influences épigénétiques qui peuvent modifier les modèles d'héritage de n'importe quel trait, entraînant une «déviation» des lois de Mendel..
L'ADN mitochondrial transmet également des informations d'une génération à l'autre, tout comme ce qui est contenu dans le noyau de toutes les cellules eucaryotes. Le génome codé dans cet ADN comprend les gènes nécessaires à la synthèse de 13 polypeptides faisant partie des sous-unités de la chaîne respiratoire mitochondriale, essentielles pour les organismes à métabolisme aérobie..
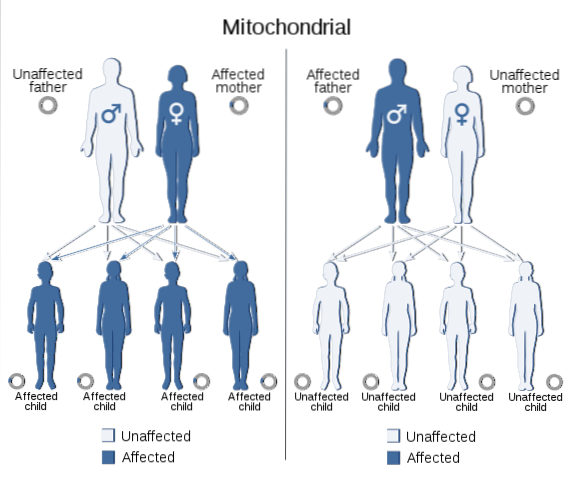
Les traits qui résultent de mutations dans le génome mitochondrial présentent un schéma de ségrégation spécifique appelé «héritage mitochondrial», qui se produit généralement par la lignée maternelle, puisque l'ovule fournit le complément total de l'ADN mitochondrial et qu'aucune mitochondrie n'est apportée par le sperme.
L'empreinte génomique consiste en une série de «marques» épigénétiques qui caractérisent certains gènes ou régions génomiques complètes et qui résultent du transit génomique du mâle ou de la femelle à travers le processus de gamétogenèse..
Il existe des grappes d'empreintes génétiques, constituées de 3 à 12 gènes répartis entre 20 et 3 700 kilos de bases d'ADN. Chaque cluster a une région connue sous le nom de région de contrôle d'impression, qui présente des modifications épigénétiques spécifiques de chaque parent, notamment:
- Méthylation de l'ADN sur des allèles spécifiques dans les résidus cytokines des paires CpG
- Modification post-traductionnelle des histones liées à la chromatine (méthylation, acétylation, phosphorylation, etc., des queues d'acides aminés de ces protéines).
Les deux types de «marques» modulent en permanence l'expression des gènes sur lesquels ils se trouvent, modifiant leurs modes de transmission à la génération suivante..
Les modèles d'hérédité dans lesquels l'expression d'une maladie dépend des allèles spécifiques hérités de l'un ou l'autre des parents sont connus sous le nom d'effet d'origine parentale..
Ce phénomène est une exception à la première loi de Mendel, qui stipule que seul un des deux allèles présents chez chaque parent est transmis à la progéniture et, selon les lois chromosomiques de l'hérédité, un seul des chromosomes homologues parentaux peut être transmis. la prochaine génération.
Il s'agit d'une exception à la règle car la disomie uniparentale est l'hérédité des deux copies d'un chromosome homologue de l'un des parents. Ce type de modèle d'héritage ne montre pas toujours des défauts phénotypiques, car il maintient les caractéristiques numériques et structurelles des chromosomes diploïdes..
Ce modèle d'héritage consiste, du point de vue phénotypique, en un mélange de traits codés par un allèle qui sont combinés. En cas de dominance incomplète, les individus hétérozygotes présentent un mélange des traits des deux allèles qui les contrôlent, ce qui implique que la relation entre les phénotypes est modifiée.
Décrit les modèles héréditaires dans lesquels les deux allèles qui sont transmis des parents à leurs enfants sont simultanément exprimés chez ceux à phénotype hétérozygote, pour lesquels les deux sont considérés comme «dominants».
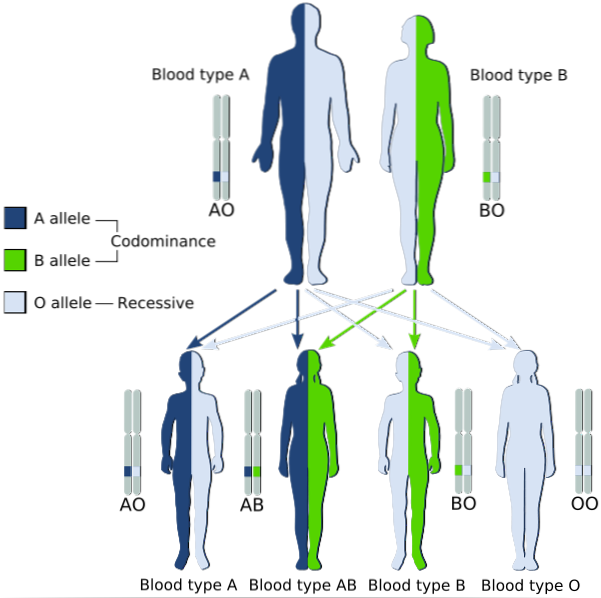
En d'autres termes, l'allèle récessif n'est pas «masqué» par l'expression de l'allèle dominant dans la paire allélique, mais les deux sont exprimés et un mélange des deux traits est observé dans le phénotype..
L'une des principales faiblesses de l'héritage mendélien est peut-être représentée par des traits codés par plus d'un allèle, ce qui est assez courant chez les humains et de nombreux autres êtres vivants..
Ce phénomène héréditaire augmente la diversité des traits codés par un gène et, en outre, ces gènes peuvent également connaître des modèles de dominance et de codominance incomplètes en plus d'une dominance simple ou complète..
Une autre des "pierres dans la chaussure" ou "jambes lâches" des théories héréditaires de Mendel a à voir avec ces gènes qui contrôlent l'apparition de plus d'un phénotype ou caractéristique visible, comme c'est le cas des gènes pléiotropes..
Dans ses travaux, Mendel n'a pas non plus considéré l'hérédité de certains allèles qui peuvent empêcher la survie de la progéniture lorsqu'elle est sous forme homozygote ou hétérozygote; ce sont les allèles mortels.
Les allèles létaux sont généralement liés à des mutations ou à des défauts dans les gènes strictement nécessaires à la survie, qui, lorsqu'ils sont transmis à la génération suivante (telles mutations), en fonction de l'homozygotie ou de l'hétérozygotie des individus, sont mortels.
Il existe des caractéristiques contrôlées par plus d'un gène (avec leurs allèles) et qui, de plus, sont fortement contrôlées par l'environnement. Chez l'homme, c'est extrêmement courant et c'est le cas pour des traits tels que la taille, les yeux, les cheveux et la couleur de la peau, ainsi que le risque de souffrir de certaines maladies.
Chez les humains et de nombreux animaux, il existe également des traits qui se trouvent sur l'un des deux chromosomes sexuels et qui sont transmis par reproduction sexuée. Beaucoup de ces traits sont considérés comme «liés au sexe» lorsqu'ils sont mis en évidence chez un seul des sexes, bien que les deux soient physiquement capables d'hériter de ces traits..
La plupart des traits liés au sexe sont associés à certaines maladies et troubles récessifs.
Il existe un trouble génétique chez l'homme connu sous le nom de syndrome de Marfan, qui est causé par une mutation dans un seul gène qui affecte simultanément la croissance et le développement (hauteur, vision et fonction cardiaque, entre autres)..
Il s'agit d'un cas considéré comme un excellent exemple du modèle d'héritage non mendélien appelé pléiotropie, dans lequel un seul gène contrôle plusieurs caractéristiques.
Les troubles génétiques qui résultent de mutations dans l'ADN mitochondrial présentent un certain nombre de variations phénotypiques cliniques car ce que l'on appelle l'hétéroplasmie se produit, où différents tissus ont un pourcentage différent de génome mitochondrial mutant et, par conséquent, présentent des phénotypes différents.
Parmi ces troubles figurent les syndromes de «déplétion» mitochondriale, qui sont un groupe de troubles autosomiques récessifs caractérisés par une réduction significative du contenu en ADN mitochondrial, qui se termine par des systèmes de production d'énergie déficients dans les organes et tissus les plus touchés..
Ces syndromes peuvent être dus à des mutations du génome nucléaire qui affectent les gènes nucléaires impliqués dans la synthèse des nucléotides mitochondriaux ou dans la réplication de l'ADN mitochondrial. Les effets peuvent être mis en évidence par des myopathies, des encéphalopathies, des anomalies hépato-cérébrales ou neuro-gastro-intestinales.



Personne n'a encore commenté ce post.