
La Recombinaison génétique c'est le processus par lequel les molécules d'acide nucléique échangent des fragments, générant une nouvelle molécule. Il est très courant dans l'ADN, mais l'ARN est également un substrat pour la recombinaison. La recombinaison est, après mutation, la source la plus importante de génération de variabilité génétique.
L'ADN participe à différents processus biochimiques. Lors de la réplication, il sert de modèle pour la génération de deux nouvelles molécules d'ADN. En transcription, il permet la génération de molécules d'ARN à partir de régions spécifiques contrôlées par un promoteur.
Mais en plus de cela, l'ADN est également capable d'échanger des fragments. Par ce processus, il génère de nouvelles combinaisons qui ne sont pas le produit des deux processus précédents, ni de la fertilisation..
Tout processus de recombinaison implique la rupture et la jonction des molécules d'ADN qui participent au processus. Ce mécanisme varie en fonction du substrat de recombinaison, des enzymes qui participent au processus et du mécanisme de son exécution..
La recombinaison dépend généralement de l'existence de régions complémentaires, similaires (sinon identiques) ou homologues entre les molécules recombinantes. Dans le cas où des molécules se recombinent dans des processus non guidés par l'homologie, la recombinaison est dite non homologue.
Si l'homologie implique une région très courte présente dans les deux molécules, la recombinaison est dite spécifique au site..
Index des articles
Ce que nous appelons l'homologie dans la recombinaison ne renvoie pas nécessairement à l'origine évolutive des molécules participantes. Nous parlons davantage du degré de similitude dans la séquence nucléotidique.
Une recombinaison non réparatrice, par exemple, se produit chez les eucaryotes pendant le processus de méiose. Sans aucun doute, il ne peut y avoir d'homologie plus grande qu'entre paires de chromosomes dans la même cellule.
C'est pourquoi ils sont appelés chromosomes homologues. Cependant, il existe des cas dans lesquels l'ADN d'une cellule échange du matériel avec un ADN étranger. Ces ADN doivent être très similaires à la recombinaison, mais ils ne doivent pas nécessairement partager le même ancêtre (homologie) pour y parvenir..
Le site d'attachement et d'échange entre deux molécules d'ADN est appelé le chiasme, et le processus lui-même est appelé réticulation. Lors du croisement, un échange de bandes est vérifié entre les ADN participants..
Cela génère un cointégration, qui sont deux molécules d'ADN physiquement jointes en une seule. Lors de la «résolution» du cointégrat, deux molécules sont générées, généralement modifiées (recombinantes).
«Résoudre», dans le cadre de la recombinaison, c'est séparer les molécules d'ADN constitutives d'un cointégrat.
Dans la recombinaison site-spécifique, deux molécules d'ADN, généralement non homologues, ont une courte séquence commune aux deux. Cette séquence est la cible d'une enzyme d'épissage spécifique.
L'enzyme, capable de reconnaître cette séquence et pas une autre, la coupe en un site particulier dans les deux molécules. Avec l'aide de quelques autres facteurs, il échange les bandes d'ADN des deux molécules participantes et forme un cointégré.
C'est la base de la formation du cointégré entre le génome de la bactérie Escherichia coli et celle du bactériophage lambda. Un bactériophage est un virus qui infecte les bactéries.
La formation de ce cointégrat est réalisée par une enzyme codée dans le génome du virus: l'intégrase lambda. Il reconnaît une séquence commune appelée attP dans le génome circulaire du virus, et attB dans celui des bactéries.
En coupant les deux séquences dans les deux molécules, il génère des segments libres, échange les bandes et joint les deux génomes. Un cercle plus grand ou cointégré est alors formé.
Lors de la cointégration, le génome du virus est porté passivement par le génome bactérien, avec lequel il se réplique. Dans cet état, on dit que le virus est à l'état de provirus, et que la bactérie lui est lysogène..
Le processus inverse, c'est-à-dire la résolution du cointégration, peut prendre plusieurs générations - voire ne pas se produire. Cependant, cela est médié enzymatiquement par une autre protéine codée par le génome du virus appelée excisionase. Lorsque cela se produit, le virus se sépare du cointégré, se réactive et provoque une lyse cellulaire..
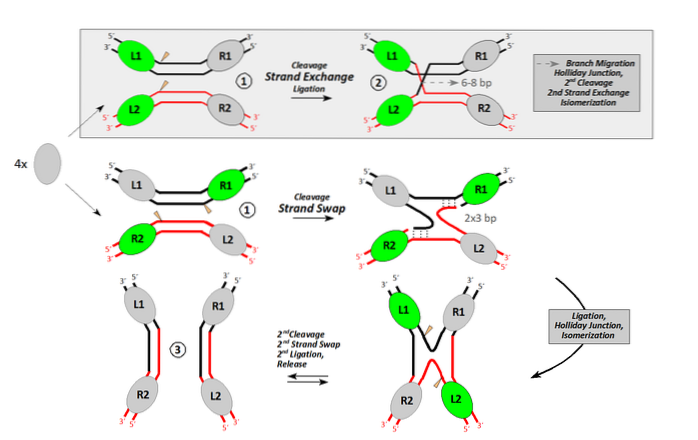
Une recombinaison homologue se produit entre des molécules d'ADN qui partagent au moins environ 40 nucléotides de similitude complète ou presque complète. Afin de réaliser le processus de recombinaison, au moins une endonucléase doit participer.
Les endonucléases sont des enzymes qui effectuent des coupes internes dans l'ADN. Certains le font pour dégrader l'ADN. D'autres, comme dans le cas de la recombinaison, le font pour générer une entaille dans l'ADN.
Ce pseudo unique permet le traitement d'un ADN à bande unique avec une extrémité libre. Cette extrémité libre, orientée par une recombinase, permet à une seule bande d'envahir un double ADN, en déplaçant la bande résidente qui lui est identique..
C'est le point de croisement, entre une molécule d'ADN donneur ("envahisseur") et un autre récepteur..
L'enzyme (recombinase) qui effectue le processus d'invasion de bande et d'échange dans Escherichia coli il s'appelle RecA. Il existe d'autres protéines homologues chez les procaryotes, comme RadA chez les archées. Chez les eucaryotes, l'enzyme équivalente est appelée RAD51.
Une fois que la bande invasive a déplacé la résidente, elle interagit avec la bande qui a été laissée simple dans la molécule donneuse. Les deux sites sont scellés par l'action d'une ligase.
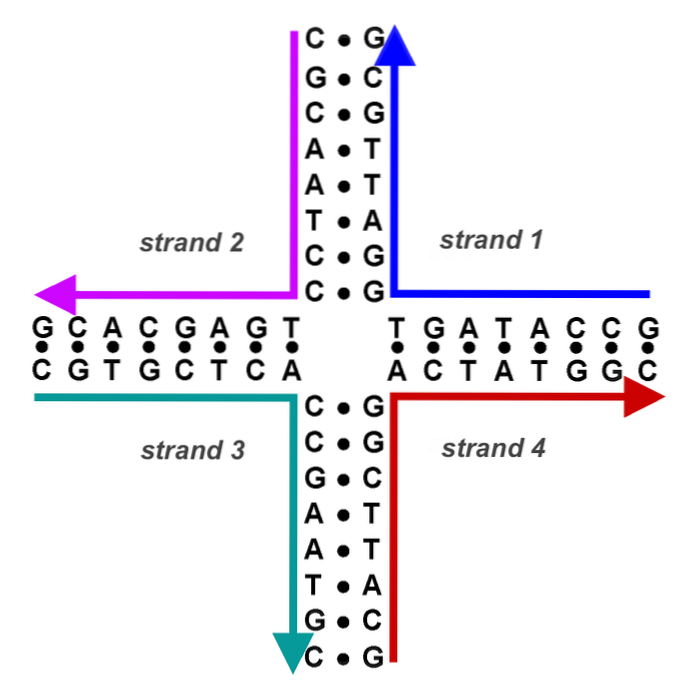
Nous avons maintenant un ADN de bandes hybrides (une bande donneuse et une bande receveuse, d'origines différentes) flanquées d'ADN donneur et d'ADN receveur. Les points de croisement (chiasmes) se déplacent dans les deux sens d'au moins 200 pb.
Chaque point de croisement forme ce que l'on appelle la structure Holliday (ADN cruciforme produit d'un événement de recombinaison).
Cet ADN cruciforme doit être résolu par d'autres endonucléases. L'ADN chimérique ou hybride de cette structure peut être résolu de deux manières. Si la deuxième coupure endonucléotidique se produit dans la même bande que la première, aucune recombinaison ne se produit. Si la deuxième coupure se produit dans l'autre bande, les produits résultants sont recombinants.
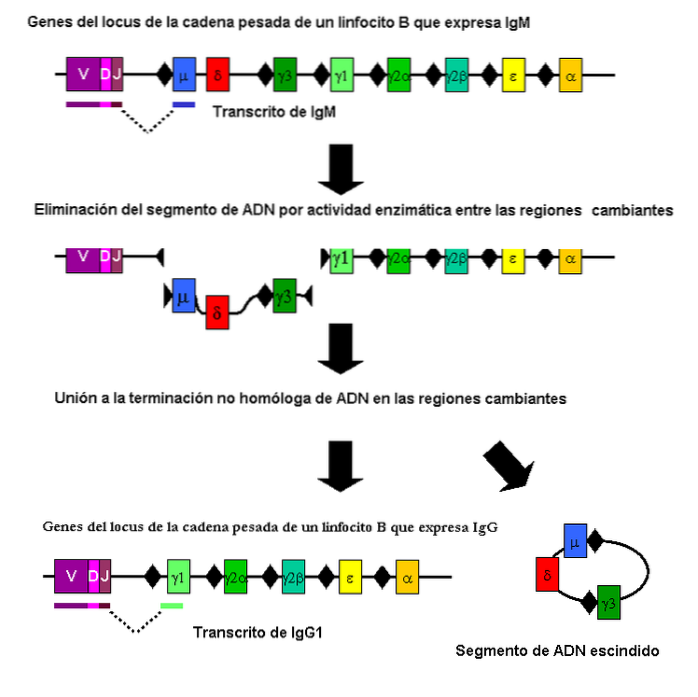
Il s'agit d'un type de recombinaison somatique (non méiotique) qui contribue à la génération de l'énorme variabilité des anticorps du système immunitaire.
Cette recombinaison a lieu notamment dans des fragments des gènes codant pour les chaînes polypeptidiques qui les définissent. Elle est réalisée par les cellules B et implique différentes régions génétiques.
Fait intéressant, il existe des parasites comme Trypanosoma brucei qui utilisent un mécanisme de recombinaison similaire pour créer une variabilité dans un antigène de surface. De cette manière, ils peuvent échapper à la réponse de l'hôte si l'hôte ne parvient pas à générer l'anticorps capable de reconnaître le «nouvel» antigène..
Enfin, il existe des processus de recombinaison qui ne dépendent pas de la similitude dans la séquence des molécules participantes. Chez les eucaryotes, par exemple, la recombinaison des extrémités non homologues est très importante.
Cela se produit avec des fragments d'ADN qui présentent des cassures de double bande dans l'ADN. Ceux-ci sont "réparés" par la cellule qui les joignent à d'autres fragments également avec des cassures de bande double.
Cependant, ces molécules ne doivent pas nécessairement être similaires pour participer à ce processus de recombinaison. Autrement dit, en réparant les dommages, la cellule peut rejoindre des ADN non liés, créant ainsi une molécule vraiment nouvelle (recombinante).
La recombinaison garantit la fidélité des informations ADN pendant et après le processus de réplication. La recombinaison détecte les dommages à l'ADN lors du nouveau processus de bandes dans cette macromolécule extrêmement longue.
Comme chaque bande a sa propre information, et celle de sa complémentarité, la recombinaison garantit qu'aucune n'est perdue. Chacun agit comme témoin de l'autre. De même, dans les organismes diploïdes, un chromosome homologue est témoin de son frère, et vice versa.
En revanche, une fois l'ADN répliqué, les mécanismes de réparation des dommages de cette molécule sont variés. Certains sont directs (la blessure est directement infligée) et d'autres sont indirects.
Les mécanismes de réparation indirecte dépendent de la recombinaison. Autrement dit, pour réparer les dommages dans une molécule d'ADN, une autre molécule homologue est utilisée. Cela agirait en recombinaison réparatrice en tant que matrice à partir de laquelle il a subi des dommages..
La recombinaison est capable de créer une énorme variabilité chromosomique au cours de la méiose. La recombinaison somatique génère également de la variabilité, comme dans le cas des anticorps chez les vertébrés.
Dans de nombreux organismes, la méiose est gamétique. Dans les organismes reproduits sexuellement, la recombinaison est l'un des moyens les plus puissants de générer de la variabilité.
Autrement dit, à la mutation spontanée et à la ségrégation chromosomique, la recombinaison doit être ajoutée comme un autre élément qui génère une variabilité gamétique..
L'intégration des génomes de bactériophages par recombinaison site-spécifique, d'autre part, a contribué au remodelage du génome de leurs bactéries hôtes..
Cela a contribué à la génération de la variabilité génomique et à l'évolution de cet important groupe d'êtres vivants..
Nous avons déjà vu que l'ADN peut être réparé, mais pas ce qui l'endommage. En réalité, presque tout peut endommager l'ADN, à commencer par une réplication défectueuse qui n'est pas corrigée..
Mais au-delà de cela, l'ADN peut être endommagé par la lumière UV, les rayonnements ionisants, les radicaux libres d'oxygène de la respiration cellulaire et ce que nous mangeons, fumons, respirons, ingérons ou touchons..
Heureusement, vous n'avez pas à renoncer à vivre pour protéger l'ADN. Certaines choses doivent être abandonnées, mais le gros travail est fait par la cellule elle-même. Ces mécanismes de détection des dommages à l'ADN, et sa réparation, ont évidemment une base génétique, et leur carence, d'énormes conséquences.
Les maladies liées à des défauts de recombinaison homologue comprennent, par exemple, les syndromes de Bloom et Werner, le cancer familial du sein et de l'ovaire, etc..



Personne n'a encore commenté ce post.