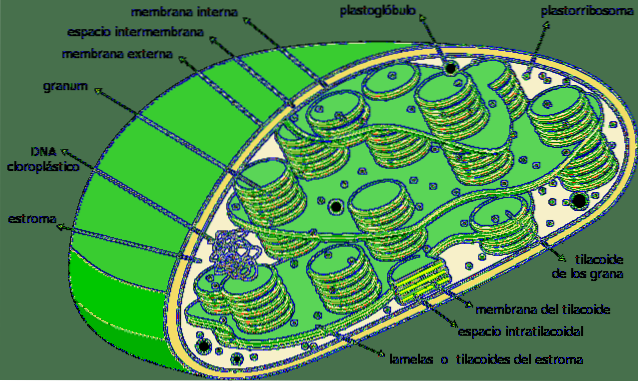
Les thylakoïdes sont des compartiments sous forme de sacs plats situés dans les chloroplastes dans les cellules végétales des plantes, dans les cyanobactéries et les algues. Ils sont généralement organisés dans une structure appelée grana -plural granum- et ça ressemble à un tas de pièces.
Les thylakoïdes sont considérés comme le troisième système membranaire des chloroplastes, à l'exception de la membrane interne et externe dudit organite. La membrane de cette structure sépare l'intérieur du thylacoïde du stroma du chloroplaste et possède une série de pigments et de protéines impliqués dans les voies métaboliques..
Dans les thylacoïdes, il existe des réactions biochimiques essentielles à la photosynthèse, un processus par lequel les plantes absorbent la lumière du soleil et la transforment en glucides. Plus précisément, ils ont la machinerie nécessaire ancrée à leur membrane pour effectuer la phase dépendante du soleil, où la lumière est piégée et convertie en énergie (ATP) et NADPH..
Index des articles
Les thylakoïdes sont un système membraneux interne tridimensionnel de chloroplastes. Les chloroplastes complètement matures ont 40 à 60 grains empilés, avec un diamètre compris entre 0,3 et 0,6 µm..
Le nombre de thylakoïdes qui composent les pousses varie considérablement: de moins de 10 sacs dans les plantes exposées à un ensoleillement suffisant, à plus de 100 thylakoïdes dans les plantes vivant dans des environnements extrêmement ombragés..
Les thylacoïdes empilés sont reliés les uns aux autres formant un compartiment continu à l'intérieur du chloroplaste. L'intérieur du thylacoïde est un compartiment assez spacieux de nature aqueuse..
La membrane thylacoïde est essentielle pour la photosynthèse, puisque la première étape du processus s'y déroule.
Les thylakoïdes sont les structures dominantes dans un chloroplaste pleinement mature. Si un chloroplaste est visualisé au microscope optique traditionnel, certaines espèces de grains peuvent être observées.
Ce sont les piles thylacoïdes; Pour cette raison, les premiers observateurs de ces structures les ont appelées "grana".
Avec l'aide du microscope électronique, l'image a pu être agrandie et il a été conclu que la nature de ces grains étaient en fait des thylakoïdes empilés.
La formation et la structure de la membrane thylacoïdienne dépendent de la formation du chloroplaste à partir d'un plaste encore indifférencié, connu sous le nom de protoplastide. La présence de lumière stimule la conversion en chloroplastes, et plus tard la formation de thylacoïdes empilés.
Dans les chloroplastes et les cyanobactéries, la membrane thylacoïde n'est pas en contact avec la partie interne de la membrane plasmique. Cependant, la formation de la membrane thylacoïde commence par l'invagination de la membrane interne..
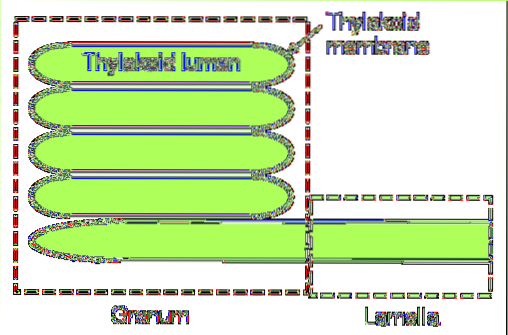
Chez les cyanobactéries et certaines espèces d'algues, les thylakoïdes sont constitués d'une seule couche de lamelles. En revanche, il existe un système plus complexe dans les chloroplastes matures..
Dans ce dernier groupe, on distingue deux parties essentielles: le grana et la lamelle du stroma. Le premier est constitué de petits disques empilés et le second est chargé de relier ces empilements les uns aux autres, formant une structure continue: la lumière du thylacoïde..
Les lipides qui composent la membrane sont hautement spécialisés et sont constitués à près de 80% de galactosyl diacylglycérol: monogalactosyl diacylglycérol et digalactosyl diacylglycérol. Ces galactolipides ont des chaînes hautement insaturées, typiques des thylakoïdes..
De même, la membrane thylacoïdienne contient moins de lipides tels que le phosphatidylglycérol. Les lipides mentionnés ne sont pas répartis de manière homogène dans les deux couches de la membrane; il existe un certain degré d'asymétrie qui semble contribuer au fonctionnement de la structure.
Les photosystèmes I et II sont les composants protéiques dominants de cette membrane. Ils sont associés au complexe cytochrome b6F et ATP synthétase.
Il a été constaté que la plupart des éléments du photosystème II sont situés dans des membranes écarlates empilées, tandis que le photosystème I est situé principalement dans des membranes thylacoïdes non empilées. Autrement dit, il existe une séparation physique entre les deux photosystèmes..
Ces complexes comprennent des protéines membranaires intégrales, des protéines périphériques, des cofacteurs et une variété de pigments..
L'intérieur du thylacoïde est constitué d'une substance épaisse et aqueuse, dont la composition est différente de celle du stroma. Participe à la photophosphorylation, en stockant les protons qui vont générer la force motrice des protons pour la synthèse de l'ATP. Dans ce processus, le pH de la lumière peut atteindre 4.
Dans le protéome de la lumière de l'organisme modèle Arabidopsis thaliana plus de 80 protéines ont été identifiées, mais leurs fonctions n'ont pas été entièrement élucidées.
Les protéines Lumen sont impliquées dans la régulation de la biogenèse des thylacoïdes et dans l'activité et le renouvellement des protéines qui forment des complexes photosynthétiques, en particulier le photosystème II et la NAD (P) H déshydrogénase..
Le processus de photosynthèse, vital pour les plantes, commence dans les thylakoïdes. La membrane qui les délimite avec le stroma chloroplastique possède toute la machinerie enzymatique nécessaire pour que les réactions photosynthétiques se produisent..
La photosynthèse peut être divisée en deux étapes majeures: les réactions lumineuses et les réactions sombres.
Comme son nom l'indique, les réactions appartenant au premier groupe ne peuvent se dérouler qu'en présence de lumière, tandis que celles du second groupe peuvent survenir avec ou sans lumière. Notez qu'il n'est pas nécessaire que l'environnement soit "sombre", il est uniquement indépendant de la lumière.
Le premier groupe de réactions, les "légères", se produit dans le thylacoïde et peut se résumer comme suit: lumière + chlorophylle + 12 HdeuxO + 12 NADP+ + 18 ADP + 18 Pje à 6 Odeux + 12 NADPH + 18 ATP.
Le deuxième groupe de réactions se produit dans le stroma chloroplastique et absorbe l'ATP et le NADPH synthétisés dans la première étape pour réduire le carbone du dioxyde de carbone en glucose (C6H12OU ALORS6). La deuxième étape peut être résumée comme suit: 12 NADPH + 18 ATP + 6 COdeux à C6H12OU ALORS6 + 12 NADP+ + 18 ADP + 18 Pje + 6 heuresdeuxOU ALORS.
Les réactions lumineuses impliquent une série de structures appelées photosystèmes, qui se trouvent dans la membrane thylacoïdienne et contiennent quelque 300 molécules de pigment, dont la chlorophylle..
Il existe deux types de photosystème: le premier a un pic d'absorption lumineuse maximal de 700 nanomètres et est connu sous le nom de P700, tandis que le second s'appelle P680. Les deux sont intégrés dans la membrane thylacoïdienne.
Le processus commence lorsque l'un des pigments absorbe un photon et que celui-ci "rebondit" vers d'autres pigments. Lorsqu'une molécule de chlorophylle absorbe la lumière, un électron saute et une autre molécule l'absorbe. La molécule qui a perdu l'électron est maintenant oxydée et a une charge négative.
Le p680 piège l'énergie lumineuse de la chlorophylle a. Dans ce photosystème, un électron est jeté dans un système d'énergie plus élevé qu'un accepteur d'électrons primaire..
Cet électron tombe dans le photosystème I en passant par la chaîne de transport d'électrons. Ce système de réactions d'oxydation et de réduction est responsable du transfert des protons et des électrons d'une molécule à une autre..
En d'autres termes, il y a un flux d'électrons de l'eau vers le photosystème II, le photosystème I et le NADPH..
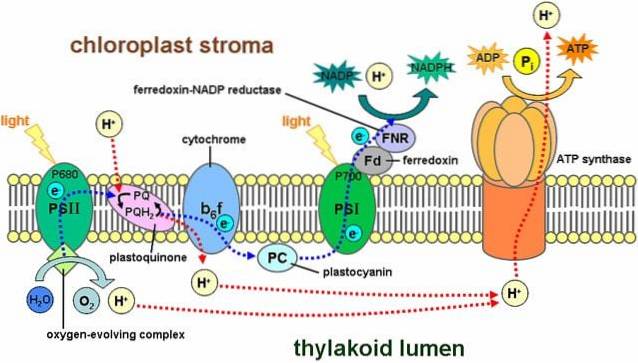
Une partie des protons générés par ce système de réaction est située à l'intérieur du thylacoïde (également appelé lumière thylacoïde), créant un gradient chimique qui génère une force motrice de protons..
Les protons se déplacent de l'espace thylacoïde vers le stroma, suivant favorablement le gradient électrochimique; c'est-à-dire qu'ils sortent du thylacoïde.
Cependant, le passage des protons ne se fait par aucun site de la membrane, ils doivent le faire à travers un système enzymatique complexe appelé ATP synthétase..
Ce mouvement des protons vers le stroma provoque la formation d'ATP à partir de l'ADP, un processus analogue à celui qui se produit dans les mitochondries. La synthèse d'ATP à l'aide de la lumière est appelée photophosphorylation.
Ces étapes mentionnées se produisent simultanément: la chlorophylle du photosystème II perd un électron et doit le remplacer par un électron issu de la décomposition d'une molécule d'eau; le photosystème I emprisonne la lumière, s'oxyde et libère un électron piégé par le NADP+.
L'électron perdu du photosystème I est remplacé par celui résultant du photosystème II. Ces composés seront utilisés dans les réactions ultérieures de fixation du carbone, dans le cycle de Calvin..
L'évolution de la photosynthèse en tant que processus de libération d'oxygène a permis la vie telle que nous la connaissons.
Il est proposé que la photosynthèse se soit développée il y a quelques milliards d'années chez l'ancêtre qui a donné naissance aux cyanobactéries actuelles, à partir d'un complexe photosynthétique anoxique.
Il est proposé que l'évolution de la photosynthèse ait été accompagnée de deux événements indispensables: la création du photosystème P680 et la genèse d'un système membranaire interne, sans connexion à la membrane cellulaire.
Il existe une protéine appelée Vipp1 essentielle à la formation des thylakoïdes. En effet, cette protéine est présente dans les plantes, les algues et les cyanobactéries, mais absente dans les bactéries qui réalisent la photosynthèse anoxique..
On pense que ce gène pourrait provenir de la duplication de gènes chez l'ancêtre possible des cyanobactéries. Il n'y a qu'un seul cas de cyanobactéries capables de photosynthèse avec l'oxygène et sans thylakoïdes: l'espèce Gloeobacter violaceus.

Personne n'a encore commenté ce post.