
le Appareil de Golgi, Aussi connu sous le nom de complexe de Golgi, il s'agit d'un organite cellulaire membraneux formé par un ensemble de vésicules plates empilées ensemble; Ces sacs contiennent du liquide à l'intérieur. Trouvé dans une grande variété d'eucaryotes, y compris les animaux, les plantes et les champignons.
Cet organite est responsable du traitement, du conditionnement, de la classification, de la distribution et de la modification des protéines. De plus, il joue également un rôle dans la synthèse des lipides et des glucides. D'autre part, dans les plantes, la synthèse des composants de la paroi cellulaire se produit dans l'appareil de Golgi..
L'appareil de Golgi a été découvert en 1888, alors que les cellules nerveuses étaient étudiées; son découvreur, Camillo Golgi, a remporté le prix Nobel. La structure est détectable par coloration au chromate d'argent.
Au début, l'existence de l'organe était douteuse pour les scientifiques de l'époque et ils attribuaient les observations de Golgi à de simples artefacts produits des techniques utilisées..
Index des articles
L'appareil de Golgi est un organite eucaryote de nature membraneuse. Il ressemble à des sacs en piles, bien que l'organisation puisse varier en fonction du type de cellule et de l'organisme. Il est responsable de la modification des protéines après traduction.
Par exemple, certains glucides peuvent être ajoutés pour former une glycoprotéine. Ce produit est emballé et distribué dans le compartiment cellulaire où il est nécessaire, tel que la membrane, les lysosomes ou les vacuoles; il peut également être envoyé à l'extérieur de la cellule. Il participe également à la synthèse de biomolécules.
Le cytosquelette (spécifiquement l'actine) détermine son emplacement, et généralement le complexe est situé dans une zone de l'intérieur de la cellule proche du noyau et du centrosome.
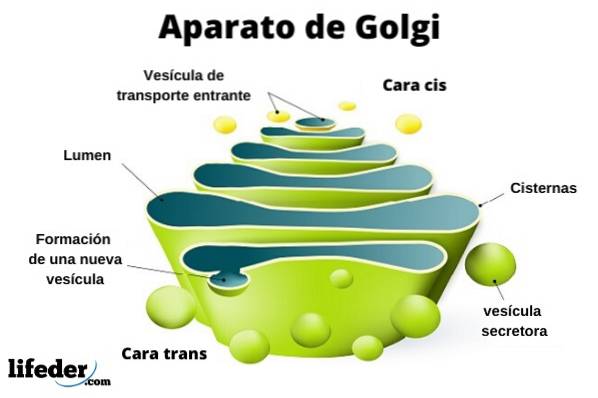
Le complexe de Golgi est constitué d'un ensemble de sacs plats, fenêtrés, en forme de disque, appelés citernes golgiennes, d'épaisseur variable.
Ces sacs sont empilés, par groupes de quatre ou six citernes. Dans une cellule de mammifère, vous pouvez trouver entre 40 et 100 batteries connectées les unes aux autres.
Le complexe de Golgi présente une caractéristique intéressante: il y a une polarité en termes de structure et aussi de fonction.
Vous pouvez faire la distinction entre le visage cis et le visage trans. Le premier est lié à l'entrée des protéines et se situe à proximité du réticulum endoplasmique. Le second est la face de sortie ou la sécrétion du produit; Ils sont formés par un ou deux réservoirs de forme tubulaire.
Parallèlement à cette structure, des vésicules constituent le système de transport. Les piles de sacs sont attachées les unes aux autres dans une structure rappelant un arc ou une date.
Chez les mammifères, le complexe de Golgi est fragmenté en plusieurs vésicules au cours des processus de division cellulaire. Les vésicules passent aux cellules filles et prennent à nouveau la forme traditionnelle du complexe.
L'organisation du complexe n'est pas commune à tous les groupes d'organismes. Dans certains types de cellules, le complexe n'est pas structuré comme des ensembles de citernes empilées en groupes; au contraire, ils sont situés individuellement. Un exemple de cette organisation est celui du champignon Saccharomyces cerevisiae.
Dans certains organismes unicellulaires, tels que le toxoplasme ou trypanosome, la présence d'un seul tas membraneux a été signalée.
Toutes ces exceptions indiquent que l'empilement des structures n'est pas indispensable pour remplir leur fonction, bien que la proximité entre les sacs rende le processus de transport beaucoup plus efficace..
De même, certains eucaryotes basaux n'ont pas ces citernes; par exemple, les champignons. Cette preuve soutient la théorie selon laquelle l'appareil est apparu dans une lignée plus tardive que les premiers eucaryotes..
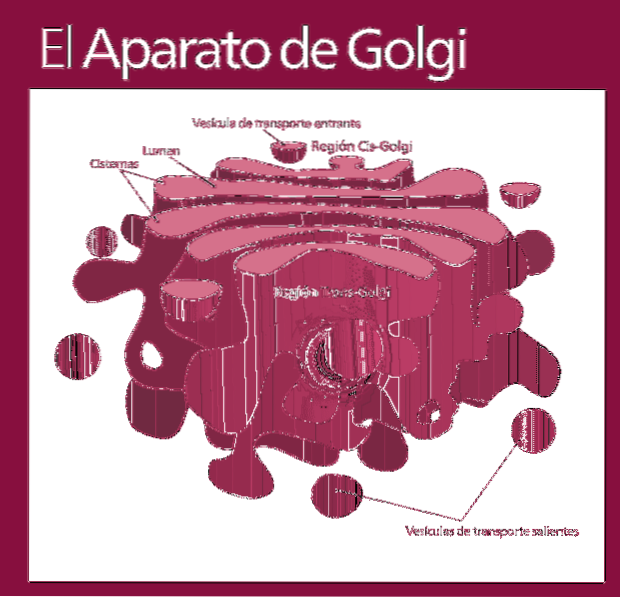
Fonctionnellement, le complexe de Golgi est divisé en les compartiments suivants: le réseau cis, les sacs empilés - qui à leur tour sont divisés en sous-compartiment intermédiaire et trans - et le trans.
Les molécules qui seront modifiées entrent dans le complexe de Golgi en suivant le même ordre (réseau cis, suivi de sous-compartiments pour être finalement excrétées dans le réseau trans).
La plupart des réactions se produisent dans les zones les plus actives: les sous-compartiments trans et intermédiaire.
Le complexe de Golgi a pour fonction principale la modification post-traductionnelle des protéines grâce aux enzymes qu'elles contiennent.
Ces modifications comprennent des processus de glycosylation (ajout de glucides), de phosphorylation (ajout d'un groupe phosphate), de sulfatation (ajout d'un groupe phosphate) et de protéolyse (dégradation des protéines).
De plus, le complexe de Golgi est impliqué dans la synthèse de biomolécules spécifiques. Chacune de ses fonctions est décrite en détail ci-dessous:
Dans l'appareil de Golgi, la modification d'une protéine en glycoprotéine se produit. Le pH acide typique de l'intérieur de l'organite est essentiel pour que ce processus se déroule normalement..
Il y a un échange constant de matériaux entre l'appareil de Golgi avec le réticulum endoplasmique et les lysosomes. Dans le réticulum endoplasmique, les protéines subissent également des modifications; ceux-ci incluent l'ajout d'un oligosaccharide.
Lorsque ces molécules (N-oligosaccharides) pénètrent dans le complexe de Golgi, elles reçoivent une série de modifications supplémentaires. Si le destin de cette molécule doit être transporté à l'extérieur de la cellule ou être reçu dans la membrane plasmique, des modifications spéciales se produisent.
Ces modifications comprennent les étapes suivantes: l'élimination de trois résidus de mannose, l'ajout de N-acétylglucosamine, l'élimination de deux mannoses et l'ajout de fucose, deux N-acétylglucosamine supplémentaires, trois galactose et trois résidus d'acide sialique..
En revanche, les protéines destinées aux lysosomes sont modifiées de la manière suivante: il n'y a pas d'élimination des mannoses dans un premier temps; au lieu de cela, la phosphorylation de ces résidus se produit. Cette étape se produit dans la région cis du complexe.
Ensuite, les groupes N-acétylglucosamine sont éliminés en laissant les mannoses avec le phosphate ajouté à l'oligosaccharide. Ces phosphates indiquent que la protéine doit être spécifiquement ciblée sur les lysosomes..
Les récepteurs responsables de la reconnaissance des phosphates qui indiquent leur destin intracellulaire sont situés dans le réseau trans.
Dans le complexe de Golgi, la synthèse des glycolipides et de la sphingomyéline se produit, en utilisant le céramide (préalablement synthétisé dans le réticulum endoplasmique) comme molécule d'origine. Ce processus est contraire à celui du reste des phospholipides qui composent la membrane plasmique, qui sont dérivés du glycérol..
La sphingomyéline est une classe de sphingolipides. C'est un composant abondant des membranes des mammifères, en particulier des cellules nerveuses, où elles font partie de la gaine de myéline.
Après leur synthèse, ils sont transportés vers leur emplacement final: la membrane plasmique. Leurs têtes polaires sont situées vers l'extérieur de la surface cellulaire; ces éléments ont un rôle spécifique dans les processus de reconnaissance cellulaire.
Dans les cellules végétales, l'appareil de Golgi contribue à la synthèse des polysaccharides qui composent la paroi cellulaire, en particulier l'hémicellulose et les pectines. Par transport vésiculaire, ces polymères sont transportés à l'extérieur de la cellule.
Chez les végétaux, cette étape est cruciale et environ 80% de l'activité du réticulum est affectée à la synthèse des polysaccharides. En fait, des centaines de ces organites ont été signalés dans des cellules végétales..
Les différentes biomolécules - protéines, glucides et lipides - sont transférées vers leurs destinations cellulaires par le complexe de Golgi. Les protéines ont une sorte de «code» qui est chargé d’informer la destination à laquelle elles appartiennent.
Ils sont transportés dans des vésicules qui quittent le réseau trans et se déplacent vers le compartiment cellulaire spécifique.
Les protéines peuvent être transportées vers la membrane par une voie constitutive spécifique. C'est pourquoi il y a une incorporation continue de protéines et de lipides dans la membrane plasmique. Les protéines dont la destination finale est le complexe de Golgi sont retenues par ce.
En plus de la voie constitutive, d'autres protéines sont destinées à l'extérieur de la cellule et se produisent à travers des signaux de l'environnement, qu'il s'agisse d'hormones, d'enzymes ou de neurotransmetteurs..
Par exemple, dans les cellules pancréatiques, les enzymes digestives sont conditionnées dans des vésicules qui ne sont sécrétées que lorsque la présence d'aliments est détectée..
Des recherches récentes rapportent l'existence de voies alternatives pour les protéines membranaires qui ne traversent pas l'appareil de Golgi. Cependant, ces voies contourne Les "non conventionnels" sont débattus dans la littérature.
Il existe cinq modèles pour expliquer le trafic de protéines dans l'appareil. Le premier concerne le trafic de matière entre les compartiments stables, chacun dispose des enzymes nécessaires pour remplir des fonctions spécifiques. Le deuxième modèle implique la maturation des citernes au fur et à mesure.
Le troisième propose également la maturation des sacs mais avec l'incorporation d'un nouveau composant: le transport tubulaire. Selon le modèle, les tubules sont importants dans le trafic dans les deux sens.
Le quatrième modèle propose que le complexe fonctionne comme une unité. Le cinquième et dernier modèle est le plus récent et maintient que le complexe est divisé en différents compartiments.
Dans certains types de cellules, le complexe de Golgi a des fonctions spécifiques. Les cellules du pancréas ont des structures spécialisées pour la sécrétion d'insuline.
Les différents groupes sanguins chez l'homme sont un exemple de modèles de glycosylation différentiels. Ce phénomène s'explique par la présence de différents allèles codant pour la glucotransférase.

Personne n'a encore commenté ce post.