La pompe sodium potassium Il s'agit d'une structure protéique incluse dans un ensemble plus large de molécules présentes dans de nombreuses membranes cellulaires, et qui sont responsables du transport actif d'ions ou d'autres petites molécules contre leurs gradients de concentration. Ils utilisent l'énergie libérée par l'hydrolyse de l'ATP et c'est pourquoi ils sont généralement appelés ATPases..
La pompe sodium potassium est une Na + / K + ATPase car elle libère l'énergie contenue dans la molécule d'ATP pour déplacer le sodium de l'intérieur vers l'extérieur de la cellule, tout en introduisant du potassium.
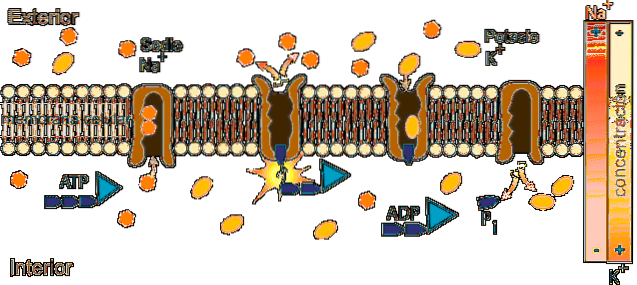
À l'intérieur de la cellule, le sodium est moins concentré (12 mEq / L) qu'à l'extérieur (142 mEq / L), tandis que le potassium est plus concentré à l'extérieur (4 mEq / L) qu'à l'intérieur (140 mEq / L).
Les pompes ATPase sont classées en trois grands groupes:
La pompe sodium potassium (Na + / K + ATPase) appartient au groupe des pompes ioniques de classe P et a été découverte en 1957 par Jens Skou, un chercheur danois, alors qu'il étudiait le mécanisme d'action des anesthésiques sur les nerfs d'un crabe. (Carcinus maenas); travail pour lequel il a reçu le prix Nobel de chimie en 1997.
Index des articles
La pompe sodium potassium est une enzyme qui du point de vue de sa structure quaternaire est formée de 2 sous-unités protéiques de type alpha (α) et deux de type beta (β).
Il s'agit donc d'un tétramère de type α2β2, dont les sous-unités sont des protéines membranaires intégrales, c'est-à-dire qu'elles traversent la bicouche lipidique et possèdent à la fois des domaines intra- et extra-cytosoliques..
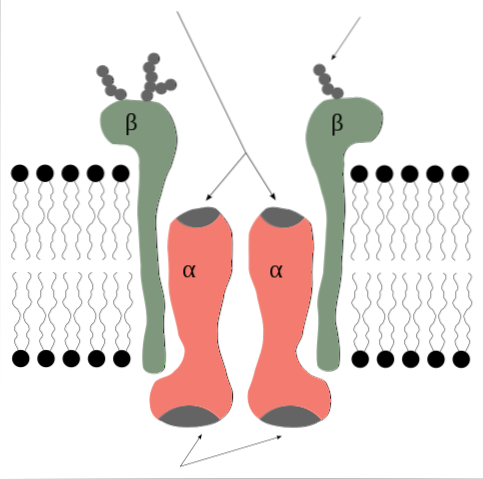
Les sous-unités α sont celles qui contiennent les sites de liaison de l'ATP et des ions Na + et K + et représentent le composant catalytique de l'enzyme et celui qui exerce la fonction de la pompe elle-même..
Les sous-unités α sont de grands polypeptides, de poids moléculaire de 120 kDa, 10 segments transmembranaires et avec leurs extrémités N- et C-terminales situées du côté cytosolique..
Du côté intracellulaire, ils ont des sites de liaison pour l'ATP et le Na +, ainsi qu'un résidu aspartate en position 376 qui représente le site qui subit le processus de phosphorylation lors de l'activation de la pompe..
Le site de liaison pour K + semble être du côté extracellulaire.
Les sous-unités β ne semblent pas avoir une participation directe à la fonction de pompage, mais en leur absence cette fonction ne se produit pas..
Les sous-unités β ont un poids moléculaire d'environ 55 kDa chacune et sont des glycoprotéines avec un seul domaine transmembranaire dont les résidus glucidiques sont insérés dans la région extracellulaire..
Ils semblent nécessaires dans le réticulum endoplasmique, où ils contribueraient au bon repliement des sous-unités α, puis, au niveau de la membrane, à stabiliser le complexe..
Les deux types de sous-unités sont hétérogènes et des isoformes α1, α2 et α3 ont été décrites pour l'une, et β1, β2 et β3 pour l'autre. Α1 se trouve dans les membranes de la plupart des cellules, tandis que α2 est présent dans les muscles, le cœur, le tissu adipeux et le cerveau et α3 dans le cœur et le cerveau.
L'isoforme β1 est celle avec la distribution la plus diffuse, bien qu'elle soit absente dans certains tissus tels que les cellules vestibulaires de l'oreille interne et les cellules musculaires glycolytiques à réponse rapide. Ces derniers ne contiennent que du β2.
Les différentes structures des sous-unités qui composent la pompe Na + / K + dans les différents tissus peuvent être dues à des spécialisations fonctionnelles qui n'ont pas encore été élucidées..
A tout instant considéré, la membrane plasmique constitue une frontière de séparation entre le compartiment correspondant à l'intérieur d'une cellule et celui qui représente le fluide extracellulaire dans lequel elle est plongée..
Les deux compartiments ont une composition qui peut être qualitativement différente, car il y a des substances à l'intérieur des cellules qui ne se trouvent pas à l'extérieur d'elles et le fluide extracellulaire contient des substances qui ne sont pas présentes au niveau intracellulaire..
Les substances présentes dans les deux compartiments peuvent être trouvées à des concentrations différentes, et ces différences peuvent avoir une signification physiologique. Tel est le cas avec de nombreux ions.
La pompe Na + / K + joue un rôle fondamental dans le maintien de l'homéostasie intracellulaire en contrôlant les concentrations d'ions sodium et potassium. Ce maintien de l'homéostasie est obtenu grâce à:
Na + a une concentration extracellulaire d'environ 142 mEq / L, tandis que sa concentration intracellulaire n'est que de 12 mEq / L; K +, en revanche, est plus concentré à l'intérieur de la cellule (140 mEq / L) qu'à l'extérieur (4 mEq / L).
Bien que la charge électrique de ces ions ne permette pas leur passage à travers la membrane, il existe des canaux ioniques qui le permettent (sélectivement), qui favorisent le mouvement si les forces qui déplacent habituellement ces ions sont également présentes..
Or, ces différences de concentration sont d'une grande importance dans le préservation de l'homéostasie de l'organisme et doit être maintenu dans une sorte d'équilibre qui, s'il était perdu, produirait d'importantes altérations organiques.
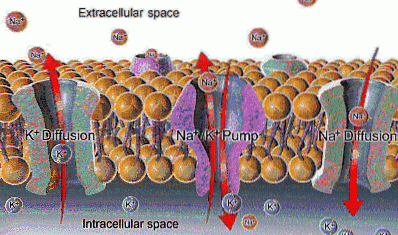
La fonction de la pompe Na + / K + permet l'extraction du sodium qui est entré par diffusion à travers des canaux ou d'autres voies de transport et la réintroduction du potassium qui s'est diffusé, permettant la conservation des concentrations intra et extracellulaires de ces ions.
Le mécanisme d'action de l'ATPase Na + / K + consiste en un cycle catalytique qui implique des réactions de transfert d'un groupe phosphoryle (Pi) et des changements conformationnels de l'enzyme qui passe d'un état E1 à un état E2 et vice versa..
L'opération nécessite la présence d'ATP et de Na + à l'intérieur de la cellule et de K + dans le fluide extracellulaire.
Le cycle commence dans l'état de conformation E1 de l'enzyme, dans lequel il y a 3 sites cytosoliques de liaison à Na + et une affinité élevée (Km de 0,6 mM) qui sont entièrement occupés car la concentration intra-ionique (12 mM) le permet.
Dans cet état (E1) et avec Na + attaché à ses sites de liaison, l'ATP se lie à son site dans le secteur cytosolique de la molécule, s'hydrolyse et un groupe phosphate est transféré à l'aspartate 376, formant un acylphosphate de haute énergie qui induit un passer à l'état E2.
Le changement conformationnel de l'état E2 implique que les sites de liaison Na + passent vers l'extérieur, leur affinité pour l'ion diminue fortement et il est libéré dans le fluide extracellulaire, tandis que, dans le même temps, l'affinité des sites de liaison K + augmente et ces ions sont attachés à l'extérieur de la pompe.


Une fois le Na + libéré et le K + lié, l'hydrolyse du phosphate d'aspartyle se produit et le changement conformationnel de l'état E2 à l'état E1 est inversé, avec la réintroduction des sites de liaison Na + vides et ceux du K + occupé.
Lorsque ce changement se produit, les sites pour Na + retrouvent leur affinité et ceux pour K + la perdent, avec laquelle le K + est libéré dans la cellule..
La pompe Na + / K + est présente dans la plupart sinon la totalité des cellules de mammifères, dans lesquelles elle revêt une importance générale en contribuant à maintenir leur osmolarité et donc leur volume.
L'entrée continue d'ions sodium dans la cellule détermine une augmentation du nombre intracellulaire de particules osmotiquement actives, ce qui induit l'entrée d'eau et l'augmentation de volume qui finirait par provoquer la rupture de la membrane et l'effondrement de la cellule..
Comme ces pompes n'introduisent que 2 K + pour 3 Na + qu'elles éliminent, elles se comportent électrogéniquement, ce qui signifie qu'elles «décompensent» les charges électriques internes, favorisant la production du potentiel membranaire caractéristique des cellules corporelles..
Son importance est également évidente en ce qui concerne les cellules qui composent les tissus excitables, dans lesquels les potentiels d'action sont caractérisés par l'entrée de l'ion Na +, qui dépolarise la cellule, et la sortie de K +, qui la repolarise..
Ces mouvements ioniques sont possibles grâce au fonctionnement des pompes Na + / K +, qui contribuent à la production des gradients chimiques qui déplacent les ions impliqués..
Sans ces pompes, fonctionnant en sens inverse, les gradients de concentration de ces ions se dissiperaient et l'activité excitatrice disparaîtrait..
Un autre aspect qui met en évidence l'extrême importance des pompes sodium-potassium est lié à la fonction rénale, ce qui serait impossible sans elles..
La fonction rénale implique la filtration quotidienne de plus ou moins 180 litres de plasma et de grandes quantités de substances, dont certaines doivent être excrétées, mais beaucoup doivent être réabsorbées pour ne pas être perdues dans l'urine..
La réabsorption du sodium, de l'eau et de nombreuses substances filtrées dépend de ces pompes, qui sont situées dans la membrane basolatérale des cellules qui composent les épithéliums des différents segments tubulaires des néphrons rénaux..
Les cellules épithéliales qui tapissent les tubules rénaux ont une face qui est en contact avec la lumière du tubule et s'appelle la face apicale, et une autre qui est en contact avec l'interstitium autour du tubule et appelée basolatérale..
L'eau et les substances réabsorbées doivent d'abord passer dans la cellule par l'apex puis dans l'interstitium par le basolatéral.
La réabsorption du Na + est essentielle à la fois par rapport à elle, et par rapport à celle de l'eau et à celle des autres substances qui en dépendent. L'entrée apicale de Na + dans la cellule nécessite qu'il y ait un gradient qui la déplace et qui implique une très faible concentration de l'ion à l'intérieur de la cellule..
Cette faible concentration intracellulaire de Na + est produite par les pompes à sodium de la membrane basolatérale qui travaillent de manière intensive pour éliminer l'ion des cellules dans l'interstitium..



Personne n'a encore commenté ce post.