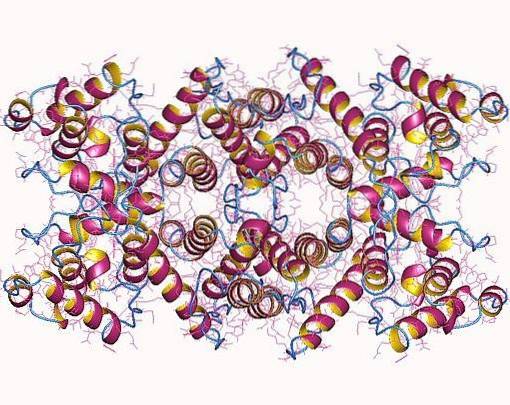
La dystrophine est une protéine en forme de bâtonnet ou de bâtonnet associée à la membrane des cellules musculaires squelettiques, lisses et cardiaques, également présente dans les cellules nerveuses et dans d'autres organes du corps humain.
Il a des fonctions similaires à celles d'autres protéines du cytosquelette et on pense qu'il agit principalement sur la stabilité de la membrane des fibres musculaires et la liaison de la membrane basale extracellulaire avec le cytosquelette intracellulaire..
Il est codé sur le chromosome X, dans l'un des plus gros gènes décrits pour l'homme, dont certaines mutations sont impliquées dans des pathologies liées aux chromosomes sexuels, comme la dystrophie musculaire de Duchenne (DMD).
Cette pathologie est le deuxième trouble héréditaire le plus répandu dans le monde. Elle affecte un homme sur 3500 et elle devient évidente entre 3 et 5 ans sous la forme d'une fonte musculaire accélérée qui peut réduire la durée de vie à 20 ans maximum..
Le gène de la dystrophine a été isolé pour la première fois en 1986 et a été caractérisé en utilisant le clonage positionnel, ce qui signifiait une grande avancée pour la génétique moléculaire de l'époque..
Index des articles
La dystrophine est une protéine très diversifiée qui est associée à la membrane plasmique des cellules musculaires (sarcolemme) et à celle d'autres cellules dans différents systèmes corporels.
Sa diversité est due aux processus liés à la régulation de l'expression du gène qui le code, qui est l'un des plus grands gènes décrits pour l'homme. En effet, il compte plus de 2,5 millions de paires de bases, ce qui représente environ 0,1% du génome..
Ce gène est principalement exprimé dans les cellules musculaires squelettiques et cardiaques ainsi que dans le cerveau, bien que dans une bien moindre mesure. Il est composé d'environ 99% d'introns et la région codante n'est représentée que dans 86 exons.
Trois isoformes différentes de cette protéine sont reconnues qui proviennent de la traduction de messagers qui sont transcrits à partir de trois promoteurs différents: une qui ne se trouve que dans les neurones corticaux et hippocampiques, une autre dans les cellules de Purkinje (également dans le cerveau), et cette dernière dans le muscle cellules (squelettiques et cardiaques).
Etant donné que le gène de la dystrophine peut être "lu" à partir de différents promoteurs internes, il existe différentes isoformes de cette protéine qui sont, bien entendu, de tailles différentes. Sur cette base, la structure des isoformes «complètes» et «courtes» est décrite ci-dessous..
Les isoformes «entières» de la dystrophine sont des protéines en forme de bâtonnet qui possèdent quatre domaines essentiels (N-terminal, domaine central, domaine riche en cystéine et domaine C-terminal) qui, ensemble, pèsent un peu plus de 420 kDa et représentent environ 3685 résidus d'acides aminés..
Le domaine N-terminal est similaire à l'α-actinine (une protéine de liaison à l'actine) et peut comprendre entre 232 et 240 acides aminés, selon l'isoforme. Le domaine de noyau ou de tige est composé de 25 répétitions triples hélicoïdales semblables à la spectrine et a environ 3000 résidus d'acides aminés.
La région C-terminale du domaine central, qui est constituée d'un domaine riche en répétitions de cystéine, a environ 280 résidus et est très similaire au motif de liaison au calcium présent dans des protéines telles que la calmoduline, l'α-actinine et β. -spectrine. Le domaine C-terminal de la protéine est composé de 420 acides aminés.
Puisque le gène de la dystrophine a au moins quatre promoteurs internes, il peut y avoir des protéines de différentes longueurs, qui diffèrent les unes des autres en raison de l'absence de l'un de leurs domaines..
Chacun des promoteurs internes a un premier exon unique qui est séparé en exons 30, 45, 56 et 63, générant des produits de 260 kDa (Dp260), 140 kDa (Dp140), 116 kDa (Dp116) et 71 kDa (Dp71), qui s'expriment dans différentes régions du corps.
Dp260 est exprimé dans la rétine et coexiste avec des formes cérébrales et musculaires «pleines». Dp140 se trouve dans le cerveau, la rétine et les reins, tandis que Dp116 ne se trouve que dans les nerfs périphériques adultes et Dp71 se trouve dans la plupart des tissus non musculaires.
Selon divers auteurs, la dystrophine a diverses fonctions qui n'impliquent pas seulement sa participation en tant que protéine du cytosquelette.
La fonction principale de la dystrophine, en tant que molécule associée à la membrane des cellules nerveuses et musculaires, est d'interagir avec au moins six protéines membranaires intégrales différentes, avec lesquelles elle se lie pour former des complexes dystrophine-glycoprotéine..
La formation de ce complexe génère un «pont» à travers la membrane des cellules musculaires ou sarcolemme et relie «de manière flexible» la lame basale de la matrice extracellulaire au cytosquelette interne.
Le complexe dystrophine-glycoprotéine fonctionne dans la stabilisation de la membrane et dans la protection des fibres musculaires contre la nécrose ou les dommages causés par la contraction induite pendant de longues périodes de temps, ce qui a été démontré par la génétique inverse..
Cette «stabilisation» est souvent considérée comme analogue à ce qu'une protéine similaire connue sous le nom de spectrine fournit des cellules telles que les érythrocytes circulant dans le sang lorsqu'ils passent à travers des capillaires étroits..
La dystrophine ou, plutôt, le complexe protéique qu'elle forme avec les glycoprotéines de la membrane a non seulement des fonctions structurelles, mais il a également été souligné qu'elle peut avoir certaines fonctions dans la signalisation et la communication cellulaires..
Son emplacement suggère qu'il peut participer à la transmission de la tension des filaments d'actine dans les sarcomères des fibres musculaires à travers la membrane plasmique vers la matrice extracellulaire, puisque celle-ci est physiquement associée à ces filaments et à l'espace extracellulaire..
Des preuves d'autres fonctions dans la transduction du signal ont émergé de certaines études menées avec des mutants pour le gène de la dystrophine, dans lesquelles des défauts dans les cascades de signalisation qui ont à voir avec la mort cellulaire programmée ou la défense cellulaire sont observés..


Personne n'a encore commenté ce post.