
La glycolyse ou la glycolyse est la principale voie de catabolisme du glucose, dont l'objectif final est d'obtenir de l'énergie sous forme d'ATP et un pouvoir réducteur sous forme de NADH, à partir de ce glucide.
Cette voie, pleinement élucidée dans les années 1930 par Gustav Embden et Otto Meyerhof lors de l'étude de la consommation de glucose dans les cellules musculaires squelettiques, consiste en l'oxydation complète de ce monosaccharide et, à elle seule, représente une voie anaérobie du glucose..
C'est l'une des principales voies métaboliques, car il se produit, avec ses différences, dans tous les organismes vivants existants, unicellulaire ou multicellulaire, procaryote ou eucaryote, et on pense qu'il s'agit d'une chaîne de réactions qui est évolutivement hautement conservée dans la nature..
En fait, certains organismes et types de cellules dépendent exclusivement de cette voie pour survivre..
Dans un premier temps, la glycolyse consiste en l'oxydation du glucose, de 6 atomes de carbone, en pyruvate, qui a trois atomes de carbone; avec la production concomitante d'ATP et de NADH, utile pour les cellules du point de vue métabolique et synthétique.
Dans les cellules capables de traiter davantage les produits obtenus à partir du catabolisme du glucose, la glycolyse se termine par la production de dioxyde de carbone et d'eau à travers le cycle de Krebs et la chaîne de transport d'électrons (glycolyse aérobie)..
Dix réactions enzymatiques ont lieu au cours de la voie glycolytique et, bien que la régulation de ces réactions puisse être quelque peu différente d'une espèce à l'autre, les mécanismes de régulation sont également assez conservés..
Index des articles
D'un point de vue métabolique, le glucose est l'un des glucides les plus importants pour tous les êtres vivants.
C'est une molécule stable et très soluble, de sorte qu'elle peut être transportée avec une relative facilité dans tout le corps d'un animal ou d'une plante, de l'endroit où elle est stockée et / ou obtenue jusqu'à l'endroit où elle est nécessaire comme carburant cellulaire..
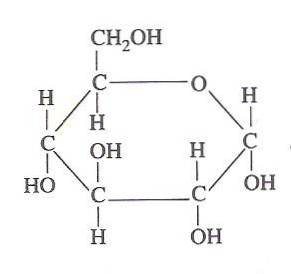
L'énergie chimique contenue dans le glucose est exploitée par les cellules vivantes grâce à la glycolyse, qui consiste en une série d'étapes hautement contrôlées par lesquelles l'énergie libérée par l'oxydation de ce glucide peut être «capturée» en des formes d'énergie plus utilisables., D'où son importance..
Par cette voie, non seulement de l'énergie (ATP) et du pouvoir réducteur (NADH) sont obtenus, mais elle fournit également une série d'intermédiaires métaboliques qui font partie d'autres voies, également importantes du fonctionnement anabolique (biosynthétique) et cellulaire général. Voici une liste:
- Glucose 6-phosphate pour la voie du pentose phosphate (PPP) Voie du pentose phosphate)
- Pyruvate pour la fermentation lactique
- Pyruvate pour la synthèse d'acides aminés (alanine, principalement)
- Pyruvate pour le cycle de l'acide tricarboxylique
- Fructose 6-phosphate, glucose 6-phosphate et dihydroxyacétone phosphate, qui fonctionnent comme des «blocs de construction» dans d'autres voies telles que la synthèse du glycogène, des acides gras, des triglycérides, des nucléotides, des acides aminés, etc..
La quantité d'ATP produite par la voie glycolytique, lorsque la cellule qui la produit ne peut pas vivre dans des conditions aérobies, est suffisante pour répondre aux besoins énergétiques d'une cellule lorsqu'elle est couplée à différents types de processus de fermentation.
Cependant, en ce qui concerne les cellules aérobies, la glycolyse sert également de source d'énergie d'urgence et sert d '«étape préparatoire» avant les réactions de phosphorylation oxydative qui caractérisent les cellules à métabolisme aérobie..
La glycolyse n'est possible que grâce à la participation des 10 enzymes catalysant les réactions qui caractérisent cette voie. Beaucoup de ces enzymes sont allostériques et changent de forme ou de conformation lorsqu'elles exercent leurs fonctions catalytiques..
Il existe des enzymes qui cassent et forment des liaisons covalentes entre leurs substrats et il y en a d'autres qui nécessitent des cofacteurs spécifiques pour remplir leurs fonctions, principalement des ions métalliques..
Structurellement parlant, toutes les enzymes glycolytiques ont un centre essentiellement stratifié. β parallèle entouré par hélices α et commandé dans plus d'un domaine. En outre, ces enzymes sont caractérisées en ce que leurs sites actifs sont généralement au niveau des sites de liaison entre les domaines..
Il est également important de noter que la régulation principale de la voie passe par le contrôle (hormonal ou métabolites) d'enzymes telles que l'hexokinase, la phosphofructokinase, la glycéraldéhyde 3-phosphate déshydrogénase et la pyruvate kinase..

La première réaction de glycolyse (phosphorylation du glucose) est catalysée par l'hexokinase (HK), dont le mécanisme d'action semble consister en un substrat de «serrage induit» qui favorise l'enzyme «lock-in» autour de l'ATP et du glucose (ses substrats) une fois s'est lié à ces.
Selon l'organisme considéré, il peut y avoir une ou plusieurs isoenzymes, dont le poids moléculaire est compris entre 50 (environ 500 acides aminés) et 100 kDa, car elles semblent se regrouper sous forme de dimères, dont la formation est favorisée par le présence de glucose, d'ions magnésium et d'ATP.
L'hexokinase a une structure tertiaire composée de feuillets alpha et bêta ouverts, bien qu'il existe de nombreuses différences structurelles dans ces enzymes.
Le glucose phosphorylé par l'hexokinase est isomérisé en fructose 6-phosphate via la phosphoglucose isomérase (PGI), également connue sous le nom de glucose 6-phosphate isomérase. L'enzyme, alors, ne supprime pas ou n'ajoute pas d'atomes, mais les réorganise au niveau structurel.
Il s'agit d'une enzyme active sous sa forme dimère (le monomère pèse plus ou moins 66 kDa) et intervient non seulement dans la glycolyse, mais aussi dans la gluconéogenèse, dans la synthèse des glucides chez les plantes, etc..
Le fructose 6-phosphate est un substrat de l'enzyme phosphofructokinase, qui est capable de re-phosphoryler cette molécule en utilisant l'ATP comme donneur de groupe phosphoryle, produisant du fructose 1,6-bisphosphate.
Cette enzyme existe chez les bactéries et les mammifères sous forme d'enzyme homotétramère (composée de quatre sous-unités identiques de 33 kDa chacune pour les bactéries et de 85 kDa chacune chez les mammifères) et dans la levure c'est un octamère (composé de sous-unités plus grandes, entre 112 et 118 kDa).
C'est une enzyme allostérique, ce qui signifie qu'elle est régulée positivement ou négativement par certains de ses produits (ADP) et par d'autres molécules comme l'ATP et le citrate..
Aussi connue sous le nom de fructose 1,6-bisphosphate aldolase, l'aldolase catalyse la dégradation catalytique du fructose 1,6-bisphosphate en dihydroxyacétone phosphate et glycéraldéhyde 3-phosphate et la réaction inverse, c'est-à-dire l'union des deux sucres pour la formation de fructose 1 , 6-bisphosphate.
En d'autres termes, cette enzyme coupe le fructose 1,6-bisphosphate de moitié, libérant deux composés C 3 phosphorylés. L'aldolase est également composée de 4 sous-unités identiques, chacune avec son propre site actif..
L'existence de deux classes (I et II) de cette enzyme a été déterminée, qui se différencient par le mécanisme de la réaction qu'elles catalysent et parce que certaines (la première) se produisent dans les bactéries et les eucaryotes «inférieurs», et les autres (la seconde ) se trouvent dans des bactéries, des protistes et des métazoaires.
L'aldolase eucaryote "supérieure" est constituée d'un homotétramère de sous-unités de poids moléculaire de 40 kDa, chacune constituée d'un fût composé de 8 feuillets β / α..
Les deux trioses phosphorylés peuvent être interconvertis l'un avec l'autre grâce à l'action de la triose-phosphate isomérase, qui permet aux deux sucres d'être utilisés tout au long de la glycolyse, garantissant la pleine utilisation de chaque molécule de glucose qui entre dans la voie..
Cette enzyme a été décrite comme l'enzyme «parfaite», car elle catalyse la réaction décrite environ un billion de fois plus rapidement qu'elle ne se produirait sans votre participation. Son site actif est au centre d'une structure bêta-baril, caractéristique de nombreuses enzymes glycolytiques..
C'est une protéine dimère, composée de deux sous-unités identiques d'environ 27 kDa, toutes deux de structure globulaire..
Le glycéraldéhyde 3-phosphate produit par l'action de l'aldolase et de la triose phosphate isomérase sert de substrat à la GAPDH, qui est une enzyme homotétramère (34-38 kDa chaque sous-unité) qui se lie en coopération à une molécule de NAD + dans chacun de ses 4 sites actifs , ainsi que 2 ions phosphate ou sulfate.
Dans cette étape de la voie, l'enzyme permet la phosphorylation de l'un de ses substrats en utilisant le phosphate inorganique comme donneur de groupe phosphoryle, avec la réduction concomitante de deux molécules NAD + et la production de 1,3-bisphosphoglycérate..
La phosphoglycérate kinase est responsable du transfert de l'un des groupes phosphate du 1,3-bisphosphoglycérate à une molécule d'ADP par phosphorylation au niveau du substrat. Cette enzyme utilise un mécanisme similaire à celui utilisé par l'hexokinase, puisqu'elle se ferme au contact de ses substrats, les protégeant des molécules d'eau interférentes..
Cette enzyme, comme d'autres qui utilisent deux substrats ou plus, a un site de liaison pour l'ADP et un autre pour le phosphate de sucre.
Contrairement aux autres enzymes décrites, cette protéine est un monomère de 44 kDa à structure bilobaire, composé de deux domaines de même taille reliés par une étroite «fente».
Le 3-phosphoglycérate subit un changement du groupe phosphate vers le carbone 2, au milieu de la molécule, qui représente un site d'instabilité stratégique qui facilite le transfert ultérieur du groupe vers une molécule d'ATP dans la dernière réaction de la voie..
Ce réarrangement est catalysé par l'enzyme phosphoglycérate mutase, une enzyme dimère pour l'homme et tétramère pour la levure, avec une taille de sous-unité proche de 27 kDa..
L'énolase catalyse la déshydratation du 2-phosphoglycérate en phosphoénolpyruvate, une étape nécessaire à la génération d'ATP dans la réaction ultérieure.
C'est une enzyme dimère composée de deux sous-unités identiques de 45 kDa. Il dépend des ions magnésium pour sa stabilité et pour le changement conformationnel nécessaire pour se lier à son substrat. C'est l'une des enzymes les plus abondamment exprimées dans le cytosol de nombreux organismes et remplit des fonctions en plus des glycolytiques..
La deuxième phosphorylation au niveau du substrat qui se produit dans la glycolyse est catalysée par la pyruvate kinase, qui est responsable du transfert du groupe phosphoryle du phosphoénolpyruvate à l'ADP et de la production de pyruvate..
Cette enzyme est plus complexe que n'importe laquelle des autres enzymes glycolytiques et chez les mammifères, c'est une enzyme homotétramère (57 kDa / sous-unité). Il existe, chez les vertébrés, au moins 4 isoenzymes: L (dans le foie), R (dans les érythrocytes), M1 (dans les muscles et le cerveau) et M2 (dans les tissus fœtaux et adultes).
La voie glycolytique se compose de dix étapes séquentielles et commence par une molécule de glucose. Au cours du processus, la molécule de glucose est "activée" ou "préparée" avec l'ajout de deux phosphates, inversant deux molécules d'ATP..
Plus tard, il est "coupé" en deux fragments et finalement il est chimiquement modifié plusieurs fois, synthétisant quatre molécules d'ATP en cours de route, de sorte que le gain net de la route correspond à deux molécules d'ATP..
De ce qui précède, on peut déduire que l'itinéraire est divisé en une phase «d'investissement» énergétique, fondamentale pour l'oxydation complète de la molécule de glucose, et une autre phase de «gain» d'énergie, où l'énergie utilisée initialement est remplacée et deux ATP nets molécules.
1- La première étape de la voie glycolytique consiste en la phosphorylation du glucose médiée par l'hexokinase (HK), pour laquelle l'enzyme utilise une molécule d'ATP pour chaque molécule de glucose phosphorylée. C'est une réaction irréversible et dépend de la présence d'ions magnésium (Mg2 +):
Glucose + ATP → Glucose 6-phosphate + ADP
2- Le glucose 6-phosphate ainsi produit est isomérisé en fructose 6-phosphate grâce à l'action de l'enzyme phosphoglucose isomérase (PGI). Il s'agit d'une réaction réversible et n'entraîne pas de dépense énergétique supplémentaire:
Glucose 6-phosphate → Fructose 6-phosphate
3- Par la suite, une autre étape d'inversion d'énergie implique la phosphorylation du fructose 6-phosphate pour former le fructose 1,6-bisphosphate. Cette réaction est catalysée par l'enzyme phosphofructokinase-1 (PFK-1). Comme la première étape de la voie, la molécule donneuse du groupe phosphate est l'ATP et c'est aussi une réaction irréversible.
Fructose 6-phosphate + ATP → Fructose 1,6-bisphosphate + ADP
4- Dans cette étape de glycolyse, il y a la décomposition catalytique du fructose 1,6-bisphosphate en dihydroxyacétone phosphate (DHAP), un cétose, et du glycéraldéhyde 3-phosphate (GAP), un aldose. Cette condensation d'aldol est catalysée par l'enzyme aldolase et est un processus réversible..
Fructose 1,6-bisphosphate → Dihydroxyacétone phosphate + glycéraldéhyde 3-phosphate
5- La dernière réaction de la phase d'inversion d'énergie consiste en l'interconversion du triose phosphate DHAP et GAP catalysée par l'enzyme triose phosphate isomérase (TIM), un fait qui ne nécessite pas d'apport énergétique supplémentaire et est également un processus réversible.
Phosphate de dihydroxyacétone ↔ Glycéraldéhyde 3-phosphate
6- Le glycéraldéhyde 3-phosphate est utilisé «en aval» dans la voie glycolytique comme substrat pour une réaction d'oxydation et un autre pour la phosphorylation, catalysée par la même enzyme, la glycéraldéhyde 3-phosphate déshydrogénase (GAPDH)..
L'enzyme catalyse l'oxydation du carbone C1 de la molécule en un acide carboxylique et sa phosphorylation dans la même position, produisant du 1,3-bisphosphoglycérate. Au cours de la réaction, 2 molécules de NAD + sont réduites pour chaque molécule de glucose et 2 molécules de phosphate inorganique sont utilisées..
2Glicéraldéhyde 3-phosphate + 2NAD + + 2Pi → 2 (1,3-bisphosphoglycérate) + 2NADH + 2H
Dans les organismes aérobies, chaque NADH ainsi produit traverse la chaîne de transport d'électrons pour servir de substrat à la synthèse de 6 molécules d'ATP par phosphorylation oxydative..
7- Il s'agit de la première étape de synthèse d'ATP dans la glycolyse et implique l'action de la phosphoglycérate kinase (PGK) sur le 1,3-bisphosphoglycérate, transférant un groupe phosphoryl (phosphorylation au niveau du substrat) de cette molécule à une molécule d'ADP, donnant 2ATP et 2 molécules de 3-phosphoglycérate (3PG) pour chaque molécule de glucose.
2 (1,3-bisphosphoglycérate) + 2ADP → 2 (3-phosphoglycérate) + 2ATP
8- Le 3-phosphoglycérate sert de substrat à l'enzyme phosphoglycérate mutase (PGM), qui le convertit en 2-phosphoglycérate par déplacement du groupe phosphoryle du carbone 3 au carbone 2 par une réaction en deux étapes réversible et dépendante de ions magnésium (Mg + 2).
2 (3-phosphoglycérate) → 2 (2-phosphoglycérate)
9- L'enzyme énolase déshydrate le 2-phosphoglycérate et produit du phosphoénolpyruvate (PEP) par une réaction qui ne nécessite pas l'ajout d'énergie supplémentaire et dont le but est de produire un composé à haute énergie, capable de donner son groupe phosphoryle dans la réaction suivante.
2 (2-phosphoglycérate) → 2 Phosphoenolpyruvate
10- Le phosphoénolpyruvate est un substrat de l'enzyme pyruvate kinase (PYK), responsable du transfert du groupement phosphoryl de cette molécule vers une molécule d'ADP, catalysant ainsi une autre réaction de phosphorylation au niveau du substrat.
Dans la réaction, 2ATP et 2 molécules de pyruvate sont produites pour chaque glucose et la présence de potassium et de magnésium sous forme ionique est nécessaire..
2Phosphoénolpyruvate + 2ADP → 2Pyruvate + 2ATP
Le rendement net de la glycolyse, de cette manière, se compose de 2ATP et 2NAD + pour chaque molécule de glucose qui entre dans la voie..
S'il s'agit de cellules à métabolisme aérobie, alors la dégradation totale d'une molécule de glucose produit entre 30 et 32 ATP à travers le cycle de Krebs et la chaîne de transport d'électrons..
La réaction générale de la glycolyse est la suivante:
Glucose + 2NAD + + 2ADP + 2Pi → 2Pyruvate + 2ATP + 2NADH + 2H+
Par conséquent, s'il est analysé brièvement, il pourrait être garanti que les principaux produits de la voie glycolytique sont le pyruvate, l'ATP, le NADH et l'H.
Cependant, le devenir métabolique de chaque intermédiaire de réaction dépend, dans une large mesure, des besoins cellulaires, c'est pourquoi tous les intermédiaires peuvent être considérés comme des produits de réaction, et ils peuvent être énumérés comme suit:
- Glucose 6-phosphate
- Fructose 6-phosphate
- Fructose 1,6-bisphosphate
- Phosphate de dihydroxyacétone et 3-phosphate de glycéraldéhyde
- 1,3-bisphosphoglycérate
- 3-phosphoglycérate et 2-phosphoglycérate
- Phosphoenolpyruvate et pyruvate
Malgré le fait que la glycolyse, à elle seule (on pourrait parler de glycolyse anaérobie), ne produit qu'environ 5% de l'ATP qui peut être extrait du catabolisme aérobie du glucose, cette voie métabolique est essentielle pour plusieurs raisons:
- Il sert de source d'énergie «rapide», en particulier dans les situations où un animal doit sortir rapidement d'un état de repos, pour lesquelles les processus d'oxydation aérobie ne seraient pas assez rapides.
- Les fibres musculaires squelettiques «blanches» du corps humain, par exemple, sont des fibres à contraction rapide et dépendent de la glycolyse anaérobie pour fonctionner..
- Lorsque, pour une raison quelconque, une cellule doit se passer de certaines de ses mitochondries (qui sont les organites qui effectuent la phosphorylation oxydative d'une partie des produits glycolytiques, entre autres), la cellule devient plus dépendante de l'énergie obtenue via glycolytique..
- De nombreuses cellules dépendent du glucose comme source d'énergie par la voie glycolytique, parmi lesquelles le globules rouges, manque d'organites internes et de cellules de l'œil (en particulier celles de la cornée) qui n'ont pas une forte densité de mitochondries.



Personne n'a encore commenté ce post.