
La héritage postmendélien Il regroupe toutes les idées et / ou œuvres issues des études publiées par le naturaliste religieux Gregorio Mendel en 1865, et qui expliquent des phénomènes ou comportements génétiques différents de ceux décrits par ce.
Gregorio Mendel est considéré comme le père de la génétique, car en étudiant les croisements entre plants de pois, il a établi les premières lois de l'héritage il y a plus de 100 ans. La première loi, appelée «loi de ségrégation», propose que les gènes puissent exister sous des formes différentes ou alternatives.

Ces formes alternatives sont appelées «allèles» et elles influencent directement les caractéristiques phénotypiques d'une espèce. Les allèles peuvent déterminer la couleur des cheveux, la couleur des yeux, le type de cheveux, etc..
Dans sa première loi, Mendel a proposé que chaque trait phénotypique transmis des parents à leurs enfants soit contrôlé par deux allèles, chacun provenant d'un parent (un de la «mère» et un du «père»).
Selon cela, lorsque les parents apportent des allèles différents, la descendance aboutit à un mélange, où l'un des allèles est dominant (celui qui est exprimé dans le phénotype) et l'autre est récessif (on ne le voit pas, ou est "latent ")..
La deuxième loi de Mendel est connue sous le nom de «loi de l'héritage indépendant», car dans ses croisements Mendel a observé que le modèle d'héritage qui caractérisait les allèles qui définissaient un trait n'influençait pas le modèle d'héritage des allèles qui définissaient un trait différent.
Index des articles
Depuis que Gregor Mendel a proposé ses fameuses lois il y a plus de 100 ans, de nouveaux mécanismes ont été élucidés pour expliquer l'héritage des caractères, dont beaucoup pourraient être considérés comme contredisant ou différant considérablement de ceux décrits par Mendel..
Un exemple des phénomènes décrits par la science «moderne», dont les progrès n'ont été possibles que grâce à l'avancement des stratégies expérimentales et des méthodes de recherche, sont l'héritage lié au sexe, la dominance incomplète et la codominance, entre autres..
L'hérédité liée au sexe concerne les gènes qui ne se trouvent que sur les chromosomes sexuels. Chez l'homme, le sexe est déterminé par les chromosomes X et Y. Les femmes ont deux copies de X (XX) et les hommes en ont une de X et une de Y (XY)..
Les gènes trouvés sur chacun de ces deux chromosomes sont ceux qui contribuent aux traits liés au sexe. Les gènes sur le chromosome X sont présents chez les hommes et les femmes, mais les gènes Y ne sont présents que chez les hommes.
L'héritage lié au sexe a été observé et décrit pour la première fois par William Bateson, l'un des premiers scientifiques à «redécouvrir» les travaux de Mendel et à en profiter pour mener de nouvelles expériences basées sur ses conclusions..
Bateson a fait des croisements expérimentaux de poules avec des plumages de différentes couleurs, certains avec des plumes sombres et lisses et d'autres avec des plumes lisses et avec un motif rayé..
À travers ces croisements, il a observé que, chez la progéniture, les mâles résultants avaient toujours un plumage avec un motif rayé.
Ce n'est qu'à la troisième génération, lorsqu'un mâle rayé a été croisé avec une poule aux plumes sombres et lisses, que des coqs au plumage noir ont été obtenus. Cette proportion résultante des croix a défié les proportions proposées par Mendel.
Après les travaux de Bateson, Thomas Henry Morgan a trouvé une explication logique aux résultats étranges obtenus par Bateson, attribuant la différence de coloration chez les poulets au fait que le motif rayé était codé par un allèle dominant trouvé sur le chromosome X..
Chez les poules, contrairement à l'homme, le sexe hétérogamétique (XY) correspond aux femelles, tandis que le sexe homogamétique correspond aux mâles (XX).
L'allèle étudié par Bateson et Morgan était donc exprimé par tous ces mâles au plumage rayé, mais lorsqu'un mâle héritait de deux chromosomes X avec l'allèle correspondant à la coloration noire, alors des mâles au plumage sombre et sans motif rayé apparaissaient..
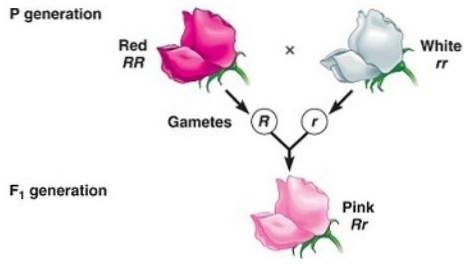
La dominance incomplète se produit lorsque le phénotype observé chez la progéniture qui résulte du croisement de deux individus homozygotes (l'un dominant et l'autre récessif) est un phénotype «intermédiaire» entre le dominant et le récessif..
Il s'agit de "l'incapacité" de l'allèle "dominant" à masquer complètement l'expression de l'allèle récessif, c'est pourquoi le phénotype résultant correspond à un mélange des traits codés par les deux allèles, souvent décrit comme un "dilué phénotype »du dominant.
Par exemple, dans les usines de Dondiego la nuit (Mirabilis jalapa) la couleur typique des pétales de fleurs sauvages est le rouge. Lorsqu'une plante à pétales rouges est croisée avec une plante de ligne pure à pétales blancs, on obtient que toute la descendance soit des pétales roses (roses).
Lorsque deux individus homozygotes dominants sont croisés et qu'un troisième phénotype est observé chez la progéniture qui correspond aux deux caractéristiques déterminées par les deux allèles parentaux, on parle de codominance.
Dans ce cas, en continuant avec un exemple similaire à celui des fleurs de l'exemple précédent, le «troisième phénotype» ne correspondrait pas à des fleurs roses, mais à une plante à fleurs blanches et fleurs rouges sur la même tige, puisque les deux allèles "codominate", et ni ne masque l'autre.
Plusieurs fois, cependant, la codominance et la dominance incomplète sont difficiles à distinguer phénotypiquement, car les deux produisent un phénotype d'un individu qui semble exprimer deux allèles différents..
Cependant, génotypiquement, les individus qui présentent des allèles codominants sont différents de ceux qui présentent une dominance incomplète, puisque dans un cas les deux allèles dominants sont exprimés dans la même quantité, et dans l'autre les proportions sont inégales..
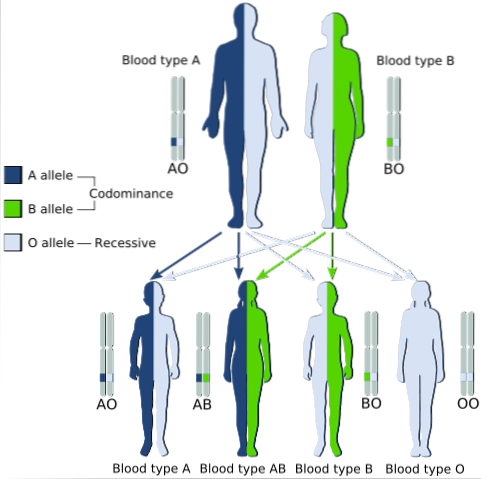
Un exemple classique de codominance est le type de groupe sanguin chez l'homme, puisque les allèles qui codent pour les antigènes A et B dans les populations humaines sont codominants..
Autrement dit, lorsque les allèles A et B sont trouvés ensemble dans les cellules d'un individu, il a le groupe sanguin AB.
Ce sont des allèles récessifs pour certains caractères qui, lorsqu'ils sont trouvés sous forme homozygote, sont mortels pour l'individu. Les individus qui présentent ces allèles ne sont pas nés, car ils empêchent le fonctionnement normal de la machinerie moléculaire et empêchent la traduction des gènes.
De nombreux endroits du génome d'un individu interagissent avec d'autres parties du même génome, c'est-à-dire que la modification spécifique d'un allèle dans le génome peut directement conditionner l'expression d'autres allèles dans les cellules d'un individu.
Ces interactions sont difficiles à déterminer et sont classées en deux types différents: les gènes qui interagissent dans la même voie génétique et ceux qui interagissent dans des voies génétiques différentes..
Un exemple classique et bien connu de gènes qui interagissent dans différentes voies génétiques concerne les gènes qui déterminent la couleur de la peau des serpents de maïs (Pantherophis guttatus), dont la couleur commune est un motif répétitif de taches noires et orange.

Ce phénotype est produit par deux pigments différents soumis à un contrôle génétique strict. Un gène détermine la couleur orange et il a deux allèles: un O +, qui code pour le pigment et un autre O-, qui détermine l'absence totale du pigment orange..
Un autre gène différent détermine la présence ou l'absence du pigment noir, les allèles de ce gène sont appelés B +, celui qui détermine la présence du pigment noir et B-, qui détermine l'absence du pigment.
Les deux gènes ne sont pas liés et déterminent indépendamment la couleur des serpents, c'est-à-dire qu'un serpent avec le génotype O + / B + est un serpent normal avec un phénotype orange et noir, mais toute variation génotypique entraînera un phénotype différent..
L'exemple classique de ces gènes est celui qui a été décrit pour les fleurs «campanulacées» (de la famille des Campanulacées), dont le phénotype «sauvage» correspond aux plantes à fleurs bleues..
La couleur bleue des fleurs de jacinthe est déterminée non pas par un gène, mais par une voie métabolique où plusieurs enzymes, chacune codée par un gène différent, participent à la synthèse d'un pigment..
Ainsi, il existe des phénotypes caractérisés en ce qu'ils sont le résultat de l'interaction de plusieurs gènes, dont les produits participent à la même voie métabolique et où la mutation ou le dysfonctionnement de l'un d'entre eux donne un phénotype, dans ce cas, de fleurs blanches dépigmentées..



Personne n'a encore commenté ce post.