le nucléosome c'est l'unité de base de l'empaquetage d'ADN dans les organismes eucaryotes. C'est donc le plus petit élément de compression de chromatine.
Le nucléosome est construit comme un octamère de protéines appelées histones, ou une structure en forme de tambour sur laquelle environ 140 nt d'ADN sont enroulés, faisant presque deux tours complets..
De plus, 40 à 80 nt supplémentaires d'ADN sont considérés comme faisant partie du nucléosome, et c'est la fraction d'ADN qui permet la continuité physique entre un nucléosome et un autre dans des structures de chromatine plus complexes (telles que la fibre de chromatine à 30 nm)..
Le code d'histone a été l'un des premiers éléments de contrôle épigénétiques à compréhension moléculaire..
Index des articles
Les nucléosomes permettent:
Dans son sens le plus élémentaire, les nucléosomes sont constitués d'ADN et de protéines. L'ADN peut être pratiquement n'importe quel ADN à double bande présent dans le noyau de la cellule eucaryote, tandis que les protéines nucléosomiques appartiennent toutes à l'ensemble des protéines appelées histones..
Les histones sont de petites protéines avec une charge élevée de résidus d'acides aminés basiques; Cela permet de contrecarrer la forte charge négative de l'ADN et d'établir une interaction physique efficace entre les deux molécules sans atteindre la rigidité de la liaison chimique covalente..
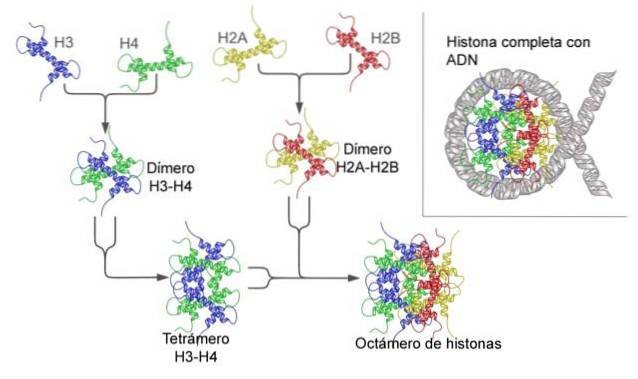
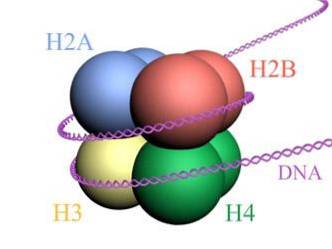
Les histones forment un octamère en forme de tambour avec deux copies ou monomères de chacune des histones H2A, H2B, H3 et H4. L'ADN fait presque deux tours complets sur les côtés de l'octamère, puis continue avec une fraction d'ADN de liaison qui s'associe à l'histone H1, pour revenir donner deux tours complets sur un autre octamère d'histone..
L'ensemble d'octamères, l'ADN associé et son ADN de liaison correspondant, est un nucléosome.
L'ADN génomique est constitué de molécules extrêmement longues (plus d'un mètre dans le cas des humains, compte tenu de tous leurs chromosomes), qui doivent être compactées et organisées au sein d'un noyau extrêmement petit.
La première étape de ce compactage est réalisée par la formation de nucléosomes. Avec cette seule étape, l'ADN est compacté environ 75 fois.
Cela donne naissance à une fibre linéaire à partir de laquelle les niveaux ultérieurs de compactage de la chromatine sont construits: la fibre à 30 nm, les boucles et les boucles de boucles..
Lorsqu'une cellule se divise, soit par mitose, soit par méiose, le degré ultime de compactage est le chromosome mitotique ou méiotique lui-même, respectivement..
Le fait que les octamères d'histones et l'ADN interagissent électrostatiquement explique en partie leur association efficace, sans perdre la fluidité nécessaire pour faire des nucléosomes des éléments dynamiques pour le compactage et la décompression de la chromatine..
Mais il y a un élément d'interaction encore plus surprenant: les extrémités N-terminales des histones sont exposées à l'extérieur de l'intérieur de l'octamère, qui est plus compact et inerte..
Ces extrémités interagissent non seulement physiquement avec l'ADN, mais subissent également une série de modifications covalentes dont dépendront le degré de compactage de la chromatine et l'expression de l'ADN associé..
L'ensemble des modifications covalentes, en termes de type et de nombre, entre autres, est collectivement connu sous le nom de code d'histone. Ces modifications comprennent la phosphorylation, la méthylation, l'acétylation, l'ubiquitination et la sumoylation des résidus arginine et lysine aux extrémités N des histones..
Chaque changement, ainsi que d'autres au sein de la même molécule ou dans les résidus d'autres histones, en particulier les histones H3, déterminera l'expression ou non de l'ADN associé, ainsi que le degré de compactage de la chromatine..
En règle générale, on a vu par exemple que les histones hyperméthylées et hypoacétylées déterminent que l'ADN associé n'est pas exprimé et que la chromatine est présente dans un état plus compact (hétérochromatique, donc inactif)..
En revanche, l'ADN euchromatique (moins compact et génétiquement actif) est associé à une chromatine dont les histones sont hyperacétylées et hypométhylées..
Nous avons déjà vu que le statut de modification covalente des histones peut déterminer le degré d'expression et le compactage local de la chromatine. Au niveau mondial, le compactage de la chromatine est également régulé par des modifications covalentes des histones dans les nucléosomes.
Il a été montré, par exemple, que l'hétérochromatine constitutive (qui n'est jamais exprimée et est densément tassée) a tendance à adhérer à la lame nucléaire, laissant les pores nucléaires libres..
De son côté, l'euchromatine constitutive (qui est toujours exprimée, comme celle qui comprend les gènes de maintenance cellulaire, et qui se situe dans des régions de chromatine laxiste), le fait en grandes boucles qui exposent l'ADN à transcrire à la machinerie de transcription..
D'autres régions de l'ADN génomique oscillent entre ces deux états en fonction du temps de développement de l'organisme, des conditions de croissance, de l'identité cellulaire, etc..
Afin de réaliser leur plan de développement, d'expression et de maintien des cellules, les génomes des organismes eucaryotes doivent régler finement quand et comment leurs potentialités génétiques doivent se manifester..
En partant alors des informations stockées dans leurs gènes, ceux-ci sont localisés dans le noyau dans des régions particulières qui déterminent leur état transcriptionnel..
On peut donc dire qu'un autre des rôles fondamentaux des nucléosomes, à travers les changements de chromatine qu'il aide à définir, est l'organisation ou l'architecture du noyau qui les abrite..
Cette architecture est héritée et est conservée phylogénétiquement grâce à l'existence de ces éléments modulaires de packaging informationnel.


Personne n'a encore commenté ce post.