le ARN ou ARN (acide ribonucléique) est un type d'acide nucléique présent dans les organismes eucaryotes et procaryotes et dans les virus. C'est un polymère nucléotidique qui contient quatre types de bases azotées dans sa structure: l'adénine, la guanine, la cytosine et l'uracile.
L'ARN se trouve généralement sous la forme d'une seule bande (sauf dans certains virus), de manière linéaire ou dans une série de structures complexes. En fait, l'ARN a un dynamisme structurel qui n'est pas observé dans la double hélice de l'ADN. Les différents types d'ARN ont des fonctions très variées.
Les ARN ribosomaux font partie des ribosomes, les structures responsables de la synthèse des protéines dans les cellules. Les ARN messagers fonctionnent comme des intermédiaires et transportent l'information génétique vers le ribosome, qui traduit le message d'une séquence nucléotidique en une séquence d'acides aminés..
Les ARN de transfert sont responsables de l'activation et du transfert des différents types d'acides aminés -20 au total vers les ribosomes. Il existe une molécule d'ARN de transfert pour chaque acide aminé qui reconnaît la séquence dans l'ARN messager.
De plus, il existe d'autres types d'ARN qui ne sont pas directement impliqués dans la synthèse des protéines et participent à la régulation des gènes..
Index des articles
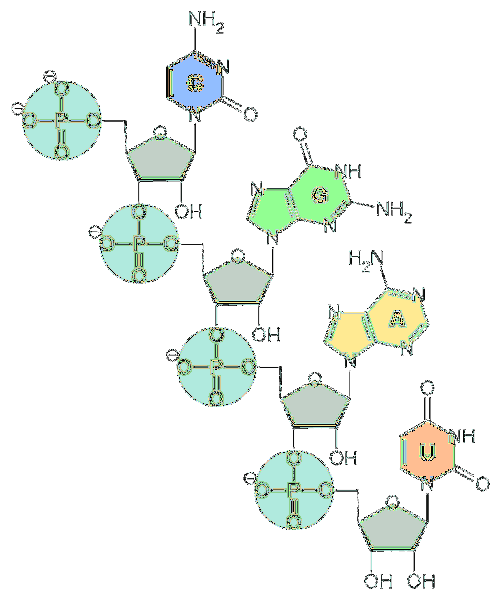
Les unités fondamentales de l'ARN sont les nucléotides. Chaque nucléotide est composé d'une base azotée (adénine, guanine, cytosine et uracile), d'un pentose et d'un groupe phosphate..
Les bases azotées sont dérivées de deux composés fondamentaux: les pyrimidines et les purines..
Les bases dérivées des purines sont l'adénine et la guanine et les bases dérivées des pyrimidines sont la cytosine et l'uracile. Bien qu'il s'agisse des bases les plus courantes, les acides nucléiques peuvent également avoir d'autres types de bases moins courants..
Quant au pentose, ce sont des unités de d-ribose. Pour cette raison, les nucléotides qui composent l'ARN sont appelés «ribonucléotides».
Les nucléotides sont liés entre eux par des liaisons chimiques impliquant le groupe phosphate. Pour les former, le groupe phosphate à l'extrémité 5 'd'un nucléotide est attaché au groupe hydroxyle (-OH) à l'extrémité 3' du nucléotide suivant, créant ainsi une liaison de type phosphodiester..
Le long de la chaîne d'acide nucléique, les liaisons phosphodiester ont la même orientation. Par conséquent, il existe une polarité du brin, distinguant les extrémités 3 'et 5'.
Par convention, la structure des acides nucléiques est représentée par l'extrémité 5 'à gauche et l'extrémité 3' à droite.
Le produit ARN de la transcription de l'ADN est une bande simple brin qui tourne vers la droite, dans une conformation hélicoïdale par l'empilement de bases. L'interaction entre les purines est beaucoup plus grande que l'interaction entre deux pyrimidines, en raison de leur taille.
Dans l'ARN, il n'est pas possible de parler d'une structure secondaire traditionnelle et de référence, telle que la double hélice d'ADN. La structure tridimensionnelle de chaque molécule d'ARN est unique et complexe, comparable à celle des protéines (logiquement, on ne peut pas globaliser la structure des protéines).
Il existe des interactions faibles qui contribuent à la stabilisation de l'ARN, en particulier l'empilement de bases, où les anneaux sont situés les uns sur les autres. Ce phénomène contribue également à la stabilité de l'hélice d'ADN..
Si la molécule d'ARN trouve une séquence complémentaire, elle pourra se coupler et former une structure double brin qui tourne vers la droite. La forme prédominante est le type A; Quant aux formes Z, elles n'ont été mises en évidence qu'en laboratoire, tandis que la forme B n'a pas été observée.
Généralement, il existe des séquences courtes (comme UUGG) qui se situent à l'extrémité de l'ARN et ont la particularité de former boucles stable. Cette séquence participe au repliement de la structure tridimensionnelle de l'ARN.
De plus, des liaisons hydrogène peuvent se former sur d'autres sites que les appariements de bases typiques (AU et CG). L'une de ces interactions se produit entre le 2'-OH du ribose avec d'autres groupes.
L'élucidation des différentes structures trouvées dans l'ARN a permis de démontrer les multiples fonctions de cet acide nucléique..
Il existe deux classes d'ARN: informatif et fonctionnel. Le premier groupe comprend les ARN qui participent à la synthèse des protéines et fonctionnent comme des intermédiaires dans le processus; les ARN informationnels sont des ARN messagers.
En revanche, les ARN appartenant à la seconde classe, les ARN fonctionnels, ne donnent pas naissance à une nouvelle molécule protéique et l'ARN lui-même est le produit final. Ce sont des ARN de transfert et des ARN ribosomiques..
Dans les cellules de mammifères, 80% de l'ARN est de l'ARN ribosomal, 15% est de l'ARN de transfert et seule une petite partie correspond à l'ARN messager. Ces trois types travaillent en coopération pour réaliser la biosynthèse des protéines..
Il existe également de petits ARN nucléaires, de petits ARN cytoplasmiques et des microARN, entre autres. Chacun des types les plus importants sera décrit en détail ci-dessous:
Chez les eucaryotes, l'ADN est confiné au noyau, tandis que la synthèse des protéines se produit dans le cytoplasme de la cellule, où se trouvent les ribosomes. En raison de cette séparation spatiale, il doit y avoir un médiateur qui transporte le message du noyau au cytoplasme et cette molécule est l'ARN messager..
L'ARN messager, ARNm abrégé, est une molécule intermédiaire qui contient les informations codées dans l'ADN et qui spécifie une séquence d'acides aminés qui donnera naissance à une protéine fonctionnelle..
Le terme ARN messager a été proposé en 1961 par François Jacob et Jacques Monod pour décrire la portion d'ARN qui transmettait le message de l'ADN aux ribosomes..
Le processus de synthèse d'un ARNm à partir du brin d'ADN est connu sous le nom de transcription et il se produit de manière différentielle entre les procaryotes et les eucaryotes..
L'expression génique est régie par plusieurs facteurs et dépend des besoins de chaque cellule. La transcription est divisée en trois étapes: l'initiation, l'élongation et la terminaison..
Le processus de réplication de l'ADN, qui se produit dans chaque division cellulaire, copie le chromosome entier. Cependant, le processus de transcription est beaucoup plus sélectif, il ne traite que du traitement de segments spécifiques du brin d'ADN et ne nécessite pas d'amorce..
Au Escherichia coli -la bactérie la mieux étudiée en sciences de la vie - la transcription commence par le déroulement de la double hélice d'ADN et la boucle de transcription est formée. L'enzyme ARN polymérase est responsable de la synthèse de l'ARN et, à mesure que la transcription se poursuit, le brin d'ADN reprend sa forme d'origine.
La transcription n'est pas initiée à des sites aléatoires sur la molécule d'ADN; il existe des sites spécialisés pour ce phénomène, appelés promoteurs. Au E. coli L'ARN polymérase est couplée à quelques paires de bases au-dessus de la région cible.
Les séquences où les facteurs de transcription sont couplés sont tout à fait conservées entre les différentes espèces. L'une des séquences promotrices les plus connues est la boîte TATA.
Lors de l'allongement, l'enzyme ARN polymérase ajoute de nouveaux nucléotides à l'extrémité 3'-OH, en suivant la direction 5 'vers 3'. Le groupe hydroxyle agit comme un nucléophile, attaquant l'alpha phosphate du nucléotide à ajouter. Cette réaction libère un pyrophosphate.
Un seul des brins d'ADN est utilisé pour synthétiser l'ARN messager, qui est copié dans la direction 3 'vers 5' (la forme antiparallèle du nouveau brin d'ARN). Le nucléotide à ajouter doit être conforme à l'appariement des bases: U paires avec A et G avec C.
L'ARN polymérase arrête le processus lorsqu'elle trouve des régions riches en cytosine et guanine. Finalement, la nouvelle molécule d'ARN messager est séparée du complexe.
Chez les procaryotes, une molécule d'ARN messager peut coder pour plus d'une protéine.
Lorsqu'un ARNm code exclusivement pour une protéine ou un polypeptide, il est appelé ARNm monocistronique, mais s'il code pour plus d'un produit protéique, l'ARNm est polycistronique (notez que dans ce contexte le terme cistron fait référence au gène).
Chez les organismes eucaryotes, la grande majorité des ARNm sont monocistroniques et la machinerie transcriptionnelle est beaucoup plus complexe dans cette lignée d'organismes. Ils se caractérisent par la présence de trois ARN polymérases, notées I, II et III, chacune avec des fonctions spécifiques.
Le I est chargé de synthétiser les pré-ARNr, le II synthétise les ARN messagers et certains ARN spéciaux. Enfin, III est responsable des ARN de transfert, du ribosomal 5S et d'autres petits ARN..
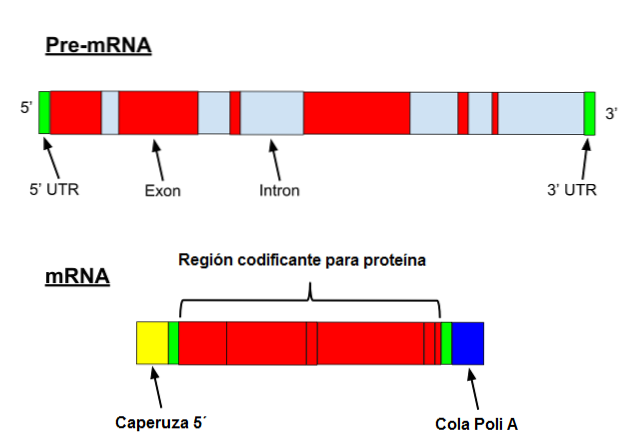
L'ARN messager subit une série de modifications spécifiques chez les eucaryotes. La première consiste à ajouter un "capuchon" à l'extrémité 5 '. Chimiquement, le capuchon est un résidu 7-méthylguanosine attaché à l'extrémité par une liaison 5 ', 5'-triphosphate..
La fonction de cette zone est de protéger l'ARN d'une éventuelle dégradation par les ribonucléases (enzymes qui décomposent l'ARN en composants plus petits).
De plus, l'élimination de l'extrémité 3 'se produit et 80 à 250 résidus d'adénine sont ajoutés. Cette structure est connue sous le nom de «queue» polyA et sert de site de liaison pour diverses protéines. Lorsqu'un procaryote acquiert une queue polyA, il a tendance à stimuler sa dégradation..
En revanche, ce messager est transcrit avec des introns. Les introns sont des séquences d'ADN qui ne font pas partie du gène mais qui «interrompent» cette séquence. Les introns ne sont pas traduits et doivent donc être supprimés du messager.
La plupart des gènes de vertébrés ont des introns, à l'exception des gènes codant pour les histones. De même, le nombre d'introns dans un gène peut varier de quelques à plusieurs dizaines d'entre eux..
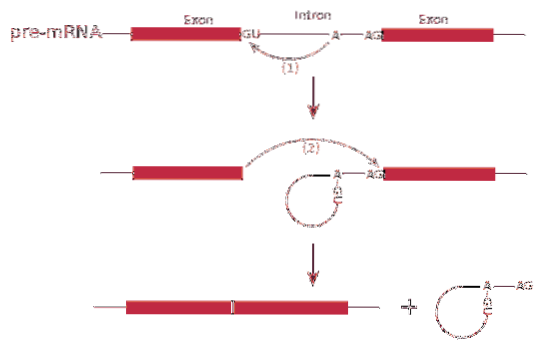
Le splissage L'ARN ou le processus d'épissage implique l'élimination des introns dans l'ARN messager.
Certains introns trouvés dans les gènes nucléaires ou mitochondriaux peuvent effectuer le processus de épissage sans l'aide d'enzymes ou d'ATP. Au lieu de cela, le processus est effectué par des réactions de transestérification. Ce mécanisme a été découvert chez le protozoaire cilié Tetrahymena thermophila.
En revanche, il existe un autre groupe de messagers qui ne sont pas capables de gérer leurs propres épissage, ils ont donc besoin de machines supplémentaires. Un assez grand nombre de gènes nucléaires appartient à ce groupe..
Le processus de épissage il est médié par un complexe protéique appelé épissosome ou complexe d'épissage. Le système est composé de complexes d'ARN spécialisés appelés petites ribonucléoprotéines nucléaires (RNP)..
Il existe cinq types de RNP: U1, U2, U4, U5 et U6, qui se trouvent dans le noyau et interviennent dans le processus de épissage.
le épissage peut produire plus d'un type de protéine - c'est ce qu'on appelle épissage alternative-, puisque les exons sont disposés de manière différentielle, créant des variétés d'ARN messagers.
L'ARN ribosomal, ARNr abrégé, se trouve dans les ribosomes et participe à la biosynthèse des protéines. C'est donc un composant essentiel de toutes les cellules.
L'ARN ribosomal s'associe à des molécules de protéines (environ 100) pour donner naissance à des présubunités ribosomales. Ils sont classés en fonction de leur coefficient de sédimentation, désigné par la lettre S pour les unités de Svedberg..
Un ribosome est composé de deux parties: la sous-unité principale et la sous-unité mineure. Les deux sous-unités diffèrent entre les procaryotes et les eucaryotes en termes de coefficient de sédimentation..
Les procaryotes ont une grande sous-unité 50S et une petite sous-unité 30S, tandis que chez les eucaryotes, la grande sous-unité est 60S et la petite 40S.
Les gènes qui codent pour les ARN ribosomiques se trouvent dans le nucléole, une zone particulière du noyau qui n'est pas délimitée par une membrane. Les ARN ribosomaux sont transcrits dans cette région par l'ARN polymérase I.
Dans les cellules qui synthétisent de grandes quantités de protéines; le nucléole est une structure proéminente. Cependant, lorsque la cellule en question ne nécessite pas un nombre élevé de produits protéiques, le nucléole est une structure presque imperceptible.
La grande sous-unité ribosomale 60S est associée aux fragments 28S et 5.8S. Concernant la petite sous-unité (40S), elle est associée à 18S.
Chez les eucaryotes supérieurs, le pré-ARNr est codé dans une unité transcriptionnelle 45S, qui implique l'ARN polymérase I. Ce transcrit est transformé en ARN ribosomaux matures 28S, 18S et 5,8S..
Au fur et à mesure que la synthèse se poursuit, le pré-ARNr s'associe à différentes protéines et forme des particules de ribonucléoprotéine. Il subit une série de modifications ultérieures qui incluent la méthylation du groupe 2'-OH du ribose et la conversion des résidus uridine en pseudouridine..
La région où ces changements se produiront est contrôlée par plus de 150 petites molécules d'ARN nucléolaire, qui ont la capacité de se coupler au pré-ARNr..
Contrairement au reste des pré-ARNr, le 5S est transcrit par l'ARN polymérase III dans le nucléoplasme et non à l'intérieur du nucléole. Après avoir été synthétisé, il est porté au nucléole pour s'assembler avec le 28S et le 5.8S, formant ainsi les unités ribosomales.
À la fin du processus d'assemblage, les sous-unités sont transférées dans le cytoplasme à travers les pores nucléaires..
Il peut arriver qu'une molécule d'ARN messager donne naissance à plusieurs protéines en même temps, se liant à plus d'un ribosome. Au fur et à mesure que le processus de traduction progresse, la fin du messager devient libre et peut être reprise par un autre ribosome, commençant une nouvelle synthèse..
Pour cette raison, il est courant de trouver des ribosomes groupés (entre 3 et 10) dans une seule molécule d'ARN messager, et ce groupe est appelé polyribosome.
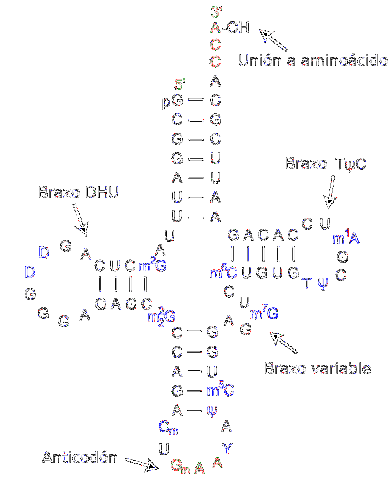
L'ARN de transfert est responsable du transfert des acides aminés à mesure que le processus de synthèse des protéines progresse. Ils sont constitués d'environ 80 nucléotides (par rapport à l'ARN messager, c'est une "petite" molécule).
La structure a des plis et des croix rappelant un trèfle à trois bras. Un anneau adénylique est situé à l'une des extrémités, où le groupe hydroxyle du ribose médie l'union avec l'acide aminé à transporter.
Les différents ARN de transfert se combinent exclusivement avec l'un des vingt acides aminés qui composent les protéines; en d'autres termes, c'est le véhicule qui transporte les éléments constitutifs des protéines. Le complexe d'ARN de transfert avec l'acide aminé est appelé aminoacyl-ARNt.
De plus, dans le processus de traduction - qui se produit grâce aux ribosomes - chaque ARN de transfert reconnaît un codon spécifique dans l'ARN messager. Une fois reconnu, l'acide aminé correspondant est libéré et fait partie du peptide synthétisé.
Pour reconnaître le type d'acide aminé qui doit être délivré, l'ARN possède un «anticodon» situé dans la région médiane de la molécule. Cet anticodon est capable de former des liaisons hydrogène avec les bases complémentaires présentes dans l'ADN messager..
Les microARN ou miARN sont un type d'ARN simple brin, entre 21 et 23 nucléotides, dont la fonction est de réguler l'expression des gènes. Comme il n'est pas traduit en protéine, il est souvent appelé ARN non codant..
Comme d'autres types d'ARN, le traitement des microARN est complexe et implique un certain nombre de protéines.
Les microARN proviennent de précurseurs plus longs appelés mi-pri-ARN, dérivés du premier transcrit du gène. Dans le noyau de la cellule, ces précurseurs sont modifiés dans le complexe microprocesseur et le résultat est un pré-miARN..
Les pré-miARN sont des épingles à cheveux de 70 nucléotides qui continuent leur traitement dans le cytoplasme par une enzyme appelée Dicer, qui assemble le complexe de silencing induit par l'ARN (RISC) et enfin le miARN est synthétisé..
Ces ARN sont capables de réguler l'expression des gènes, car ils sont complémentaires d'ARN messagers spécifiques. En s'accouplant avec leur cible, les miARN sont capables de supprimer le messager, voire de le dégrader. Par conséquent, le ribosome ne peut pas traduire ledit transcrit..
Un type particulier de microARN est les petits ARN interférents (siRNA), également appelés ARN silencing. Ce sont des ARN courts, entre 20 à 25 nucléotides, qui entravent l'expression de certains gènes.
Ce sont des instruments de recherche très prometteurs, car ils permettent de faire taire un gène d'intérêt et donc d'étudier sa fonction éventuelle..
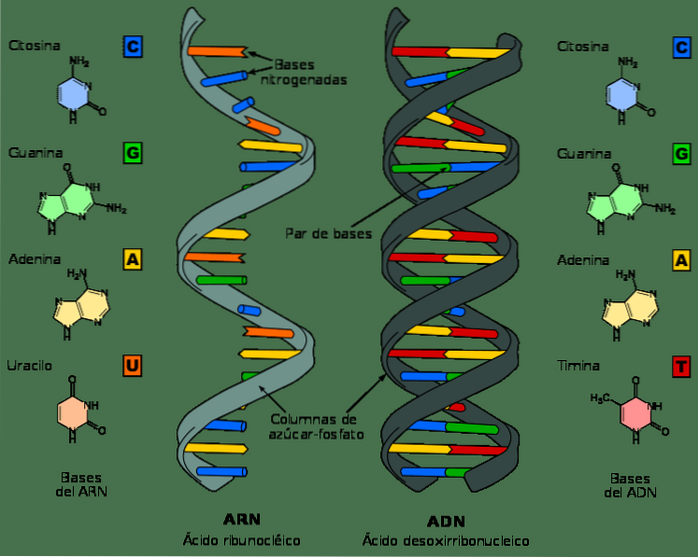
Bien que l'ADN et l'ARN soient des acides nucléiques et puissent sembler très similaires à première vue, ils diffèrent par plusieurs de leurs propriétés chimiques et structurelles. L'ADN est une molécule à double bande, tandis que l'ARN est à bande unique.
En conséquence, l'ARN est une molécule plus polyvalente et peut prendre une grande variété de formes tridimensionnelles. Cependant, certains virus ont un ARN double bande dans leur matériel génétique..
Dans les nucléotides d'ARN, la molécule de sucre est un ribose, tandis que dans l'ADN, c'est un désoxyribose, ne différant que par la présence d'un atome d'oxygène.
La liaison phosphodiester dans le squelette de l'ADN et de l'ARN est susceptible de subir un processus d'hydrolyse lent sans la présence d'enzymes. Dans des conditions alcalines, l'ARN s'hydrolyse rapidement - grâce au groupe hydroxyle supplémentaire - alors que l'ADN ne le fait pas..
De même, les bases azotées qui composent les nucléotides dans l'ADN sont la guanine, l'adénine, la thymine et la cytosine; en revanche, dans l'ARN, la thymine est remplacée par l'uracile. Uracil peut s'associer à l'adénine, de la même manière que la thymine dans l'ADN.
L'ARN est la seule molécule connue capable de stocker des informations et de catalyser des réactions chimiques en même temps; Pour cette raison, plusieurs auteurs proposent que la molécule d'ARN était cruciale dans l'origine de la vie. Étonnamment, les substrats ribosomiques sont d'autres molécules d'ARN.
La découverte des ribozymes a conduit à la redéfinition biochimique du terme «enzyme» -depuis auparavant, le terme était utilisé exclusivement pour les protéines à activité catalytique-, et a aidé à soutenir un scénario où les premières formes de vie n'utilisaient que de l'ARN comme matériel génétique.



Personne n'a encore commenté ce post.